

Nature Genetics丨群体基因组解析稻属演化历史
稻属(Oryza)涵盖27个物种及11种基因组类型,其基因组大小差异达3.4倍。该属蕴藏着近乎未开发的基因宝库,极具研究价值,为作物改良与再驯化提供了宝贵资源。
2025年4月28日,来自全球多国的研究团队,在国际顶级期刊《Nature Genetics》上发表了题为“Oryza genome evolution through a tetraploid lens”的研究论文,作者完成了11个稻属物种基因组的组装(九个四倍体、两个二倍体),揭示了核心稻属(亚)基因组仅约200Mb且高度同源,而其余核基因组组分(约80-600Mb)则呈现动态混杂、可塑性高且快速进化的特征。通过将这11个新参考基因组与既有基因组数据整合,近乎完整地揭示了稻属基因组多样化的进化机制。

一、野生稻基因组组装和TE分
作者通过PacBio和Bionano、Hi-C等测序和相关数据,完成了9个四倍体和2个二倍体野生稻种的染色体级别参考基因组组装。通过Bionano、BUSCO评估及基因组比对率等指标全面验证了其质量与连续性,并采用统一流程对基因和转座元件(TEs)进行注释。
作者分析了6个稻属特异性TE超家族的分布并构建邻接树,未发现特定TE超家族的优先扩增证据。在这些物种中,约83.4%至76.1%的LTR-RTs在多倍化事件后扩增。
作者进一步评估亚基因组TE含量差异是否源于TE积累或清除速率的差异。通过计算solo LTRs与完整LTR-RTs的比值,发现HH与JJ亚基因组间TE清除效率无显著差异,与水稻(Oryza sativa)数据一致。表明HHJJ基因组物种的亚基因组大小差异主要由HH亚基因组中LTR-RT相关序列的优先积累导致,而TE家族分布未显示特异性扩增,TE清除效率也无显著差异。
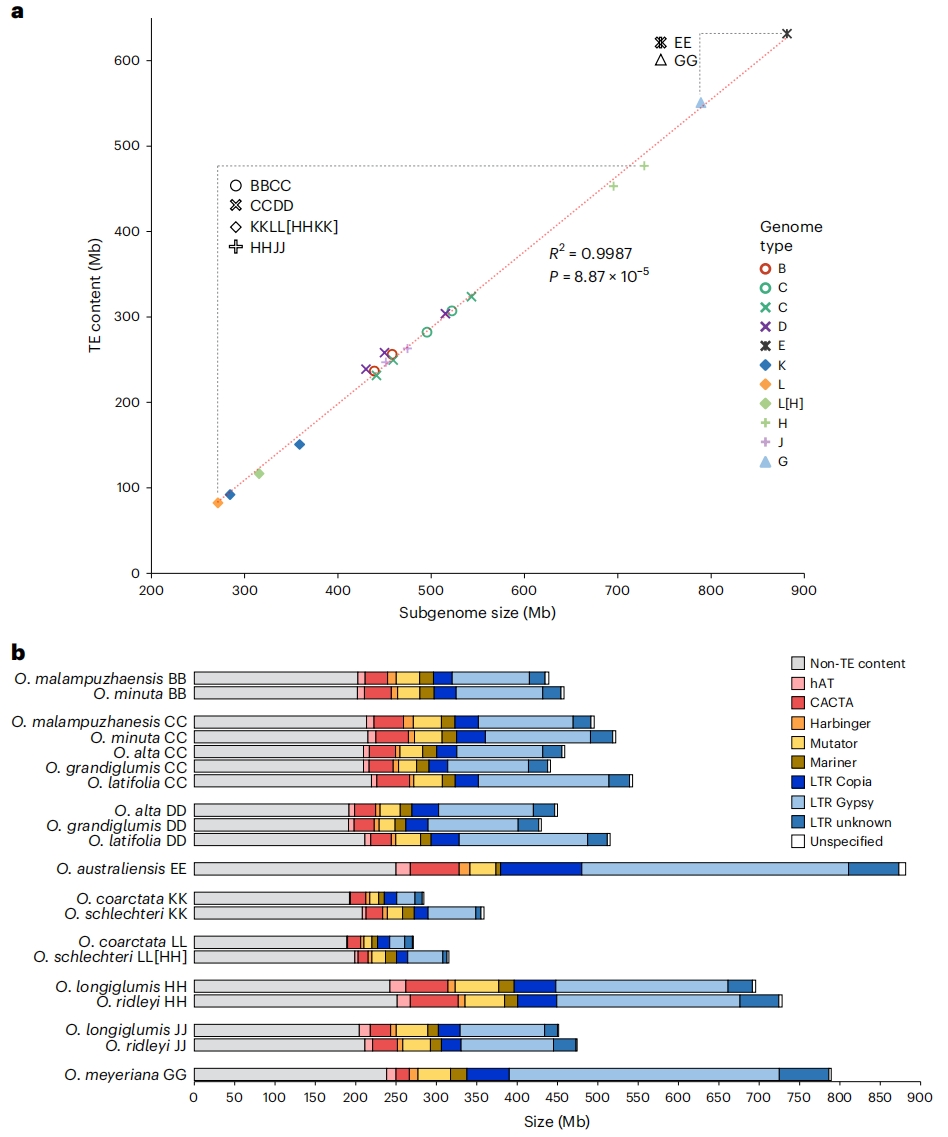
图1:稻属物种(亚)基因组大小与转座元件(TE)含量分析
二、宏观共线性与大规模染色体结构变异
为解析并直观展示稻属(Oryza)各物种间的共线性关系,作者构建了涵盖21个物种的稻属全基因组共线性图谱,以及外类群Leersia perrieri。该共线性图谱追踪了11种基因组类型(AA、BB、CC、DD、EE、KK、LL、HH、JJ、FF和GG)间的同源区块,揭示了稻属约1500万年间由倒位、重复和易位驱动的演化历程。所有稻属物种及L. perrieri均共享第11和12号染色体上的小规模片段重复。
大规模染色体结构变异常与异染色质区相关。通过对比高秆稻、大颖稻与栽培稻参考基因组,作者发现10个推定易位断点中有9个附近存在简单序列重复,提示这些序列可能促进了染色体重排。
为进一步验证上述稻属结构变异分析,作者构建了AA、BB、CC和DD基因组类型的泛基因组。各泛基因组核心区域大小为180–270 Mbp,与基于非转座子序列的估算结果高度一致。CC和DD泛基因组分析还明确支持了宏观共线性分析所检出的大规模易位,为这些重排事件提供了额外证据。
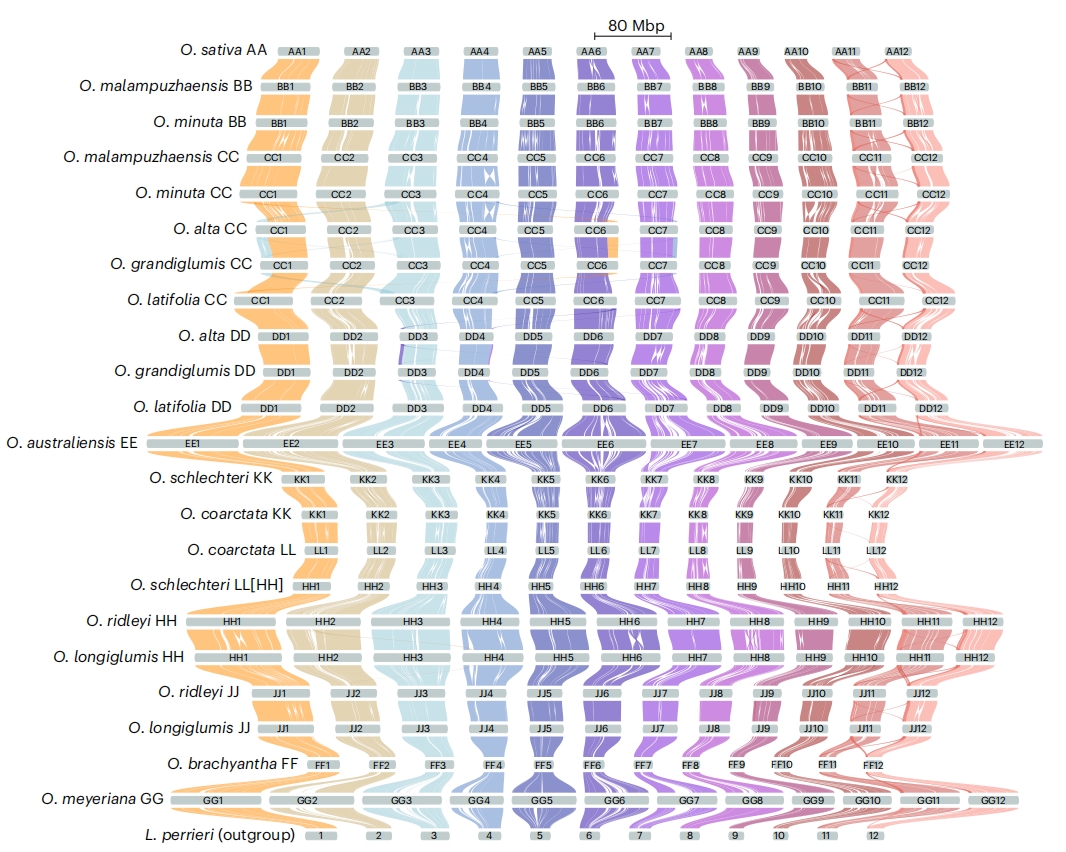
图2:12种稻属物种(21个(亚)基因组)与外类群L. perrieri的共线性图谱及大规模结构变异概览
三、共线性泛基因组分析
为鉴定稻属演化过程中保守的核心基因集以及经历复制、易位或丢失的附属基因集,作者在(亚)基因组水平进行了微观共线性分析,并通过对30个稻属(亚)基因组及外类群L. perrieri的基因预测结果构建共线性泛基因组。
共线性基因簇按分布频率可以分为四类:核心簇(8.1%,6,256个)、软核心簇(12.7%,9,865个)、非必需簇(78.9%,61,130个)和独有簇(0.3%,231个)。GO功能富集分析显示,不同类别的基因簇具有显著差异的功能特征:核心簇基因多具“必需”功能(如基础代谢、细胞维持等),而软核心簇和非必需簇基因则多具“可塑性”功能(如环境响应、次生代谢等),只在部分物种间共享。
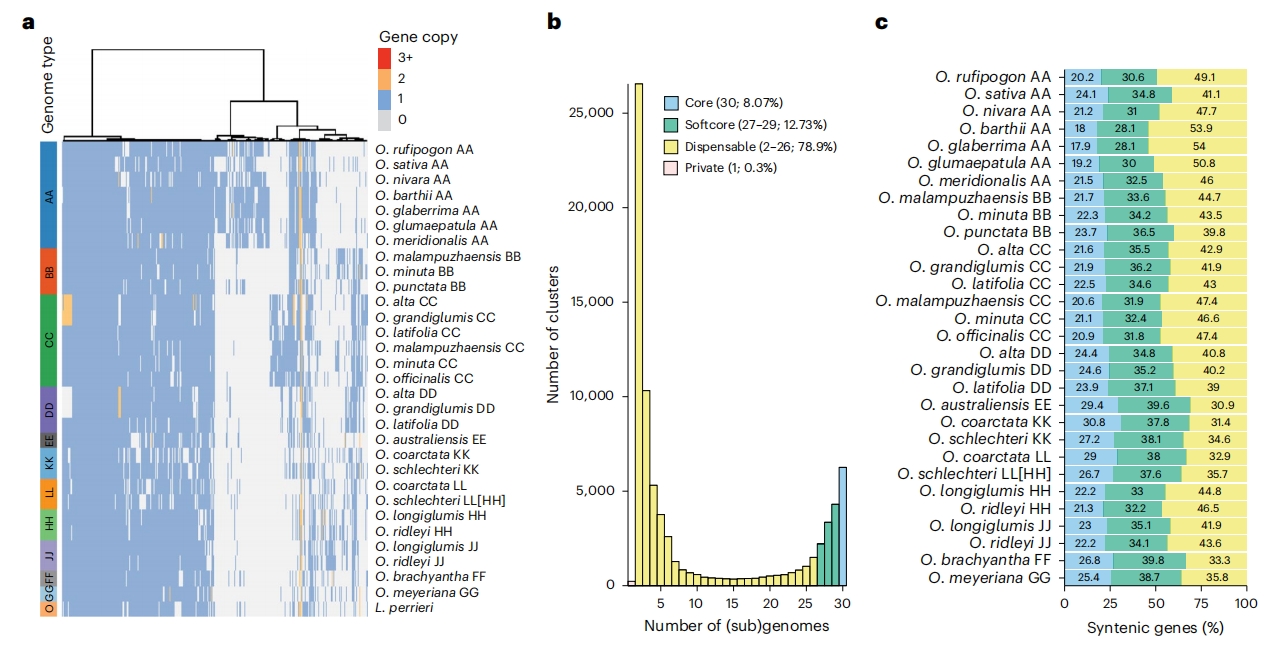
图3:共线性泛基因组构建
四、稻属演化历史的重建
为解析稻属的演化历史及母系起源,作者首先基于26个稻属物种和外类群Leersia japonica的叶绿体基因组序列构建了系统发育树。结果显示:稻属与Leersia分化后,形成基部(GG、FF和HHJJ基因型)和核心(所有基因型)两大分支,作者进一步采用3,728个单拷贝共线性基因进行溯祖系统分析。结果显示稻属(亚)基因组可分为六大单系群:AA、BB、CC、DD/EE、KK及LL/HH/JJ/FF/GG。O. coarctata的LL基因组与O. schlechteri的LL基因组聚为一支,作者通过系统发育和分子水平深入分析,最终建议将O. schlechteri的“HH”亚基因组重命名为“LL”。
为估算稻属分化时间,作者基于528个单拷贝基因的串联序列采用最小二乘定年法,所得系统树与溯祖分析结果高度一致。并推算了各基因组类型的分化时间及四倍体杂交事件发生时间。本研究显示BB与CC亚基因组先聚为一支,再与二倍体近缘种聚类,支持BBCC四倍体共同起源假说。因此,作者推定BBCC四倍体形成时间约为1.73 Myr。类似地,CCDD四倍体被认为源自单一杂交事件,其多倍化时间约为2.54 Myr。HHJJ物种,其多倍化时间约为2.24 Myr。
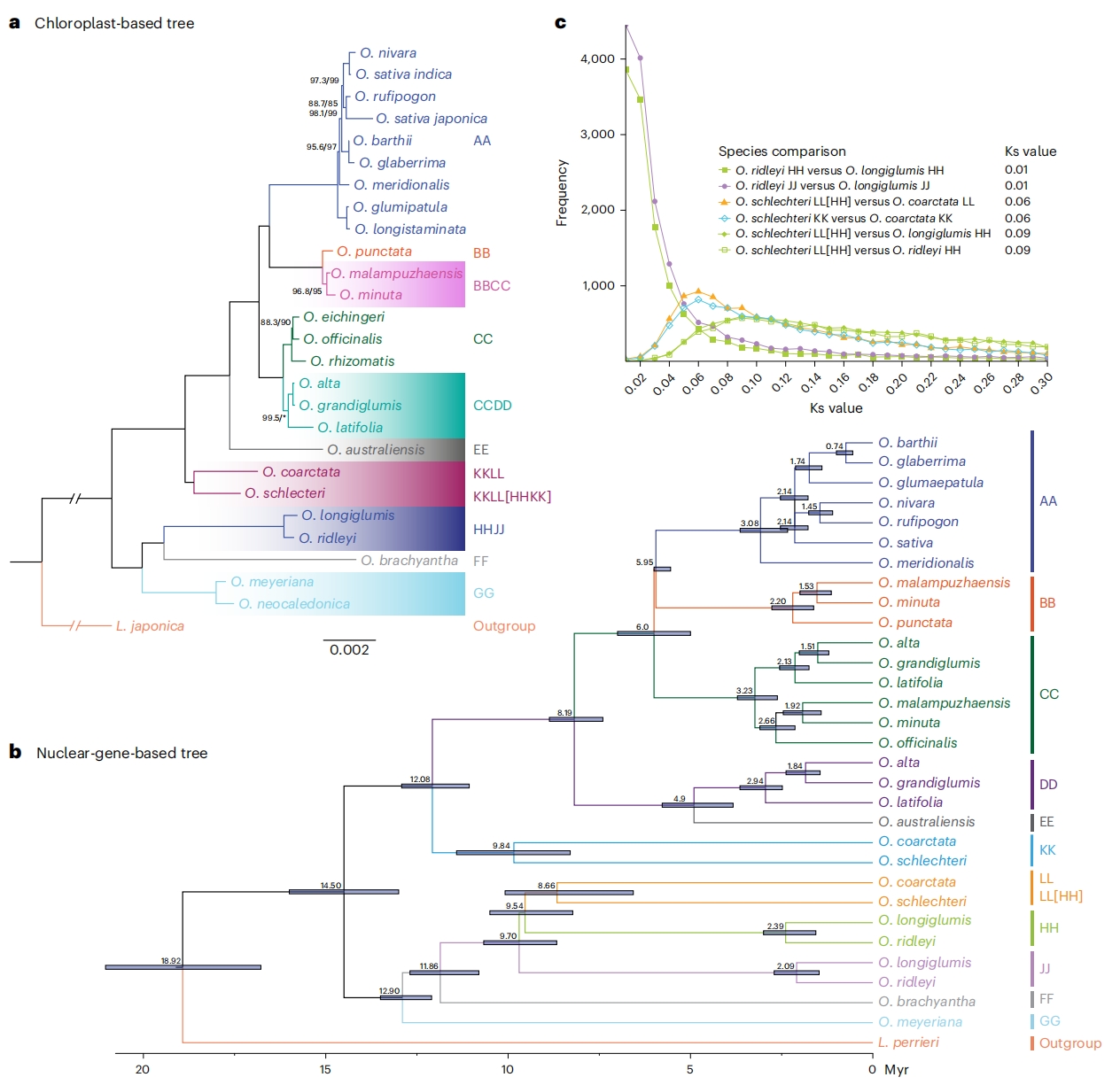
图4:稻属(亚)基因组演化研究

图5:稻属二倍体与四倍体物种起源及演化历史的系统发育共识树
五、同源基因分化现象
WGD事件发生(通过异源多倍化或同源多倍化)后,基因拷贝可能从某一同源染色体中丢失,导致基因分化。这一过程会使多倍体基因组通过基因丢失逐渐回归二倍化状态,但基因组结构已发生显著改变。作者通过统计检验显示HHJJ物种中,HH亚基因组基因保留率显著更高而药用稻复合群(officinalis complex)中,CC亚基因组基因保留率显著更高。
另外基于基因保留率平均值及多倍化时间,作者估算BBCC、CCDD和HHJJ基因组的年均基因丢失速率。结果显示较年轻物种(如BBCC)基因丢失更快,而较古老物种(如HHJJ和CCDD)基因丢失速率逐渐减缓。
六、O. coarctata中的亚基因组优势与等效性
亚基因组优势是多倍体植物中的常见现象,为探究稻属四倍体的分化模式,作者分析了耐盐野生稻O. coarctata(KKLL)的基因表达数据。结果显示平均56.1%的基因以重复拷贝形式保留在同源染色体上,其中KK亚基因组呈现过度分化(基因丢失更多)。未分化的同源基因对的表达量显著高于已分化基因。推测低表达基因在演化过程中更易丢失,这与黍属的研究一致。
另外尽管O. coarctata的LL亚基因组基因保留率更高,但同源基因表达呈现组织特异性亚基因组优势,整体表现为镶嵌式等效表达。需更多组织数据以进一步验证亚基因组等效性。
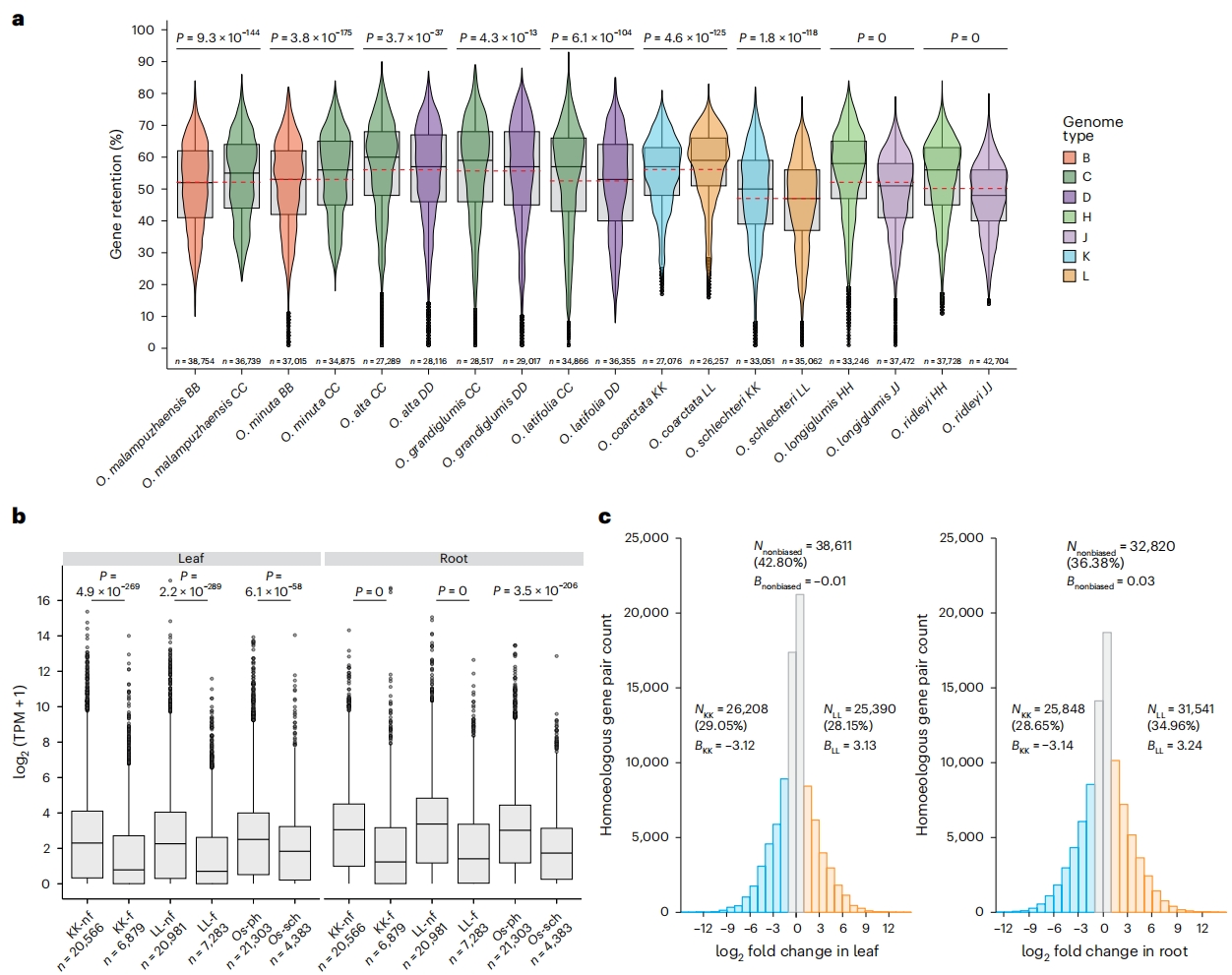
图6:稻属同源基因保留模式与O. coarctata亚基因组等效性研究
结语
本研究对9个野生四倍体稻及2个野生二倍体稻进行了染色体级别参考基因组的构建与分析。结合已发表的10个二倍体参考基因组,揭示了稻属基因组大小与组成的进化规律,发现部分物种基因组更具可塑性。基于新发现的亚基因组关系证据,作者修正了既往稻属系统发育分析。
这11个新参考基因组的发布,与既有基因组数据共同构成了一个前所未有的研究平台,将为稻属的进化生物学、功能基因组学、群体遗传学及生物多样性保护提供强大支撑,并为气候适应性水稻的再驯化奠定坚实基础。
华命生物产品服务一览

华命生物目前已开通微信公众号、抖音、知乎、B站、小红书等线上平台,欢迎感兴趣的老师扫码关注了解更多内容!
