

Nature重磅|褐藻性染色体起源与进化
性别机制在不同生物体中存在显著差异,对于两性分离的物种,性染色体携带性别决定区(SDR),并且通常在异性配子物种中不会发生重组。目前,大部分性染色体的研究集中在二倍体XX/XY和ZW/ZZ系统,而U/V单倍体性别决定系统,如苔藓和藻类,尚未得到深入探讨。在U/V系统中,性别并非在受精时决定,而是在减数分裂期间,单倍体孢子继承U或V染色体,分别发育成雌性或雄性配子体。褐藻等物种展示了多样的性别系统和生命周期,且可能具有共同的性染色体起源,这为研究提供了理想的模型。
德国马克斯-普朗克生物研究所 Susana M. Coelho研究团队于2025年8月25日在著名期刊nature ecology & evolution上发表了题为“Origin and evolutionary trajectories of brown algal sex chromosomes”的研究论文,具体分析了9种具有不同性别系统的褐藻物种的基因组,并进一步研究其性染色体进化的历史轨迹。

一、褐藻性染色体的起源
作者从数据库中下载到S. promiscuus、D. dichotoma、D. herbacea、E. crouaniorum、C. linearis、S. ischiensis、F. serratus、Ectocarpus sp. 7 以及 U. pinnatifida 的基因组,并自行完成了D. dudresnayi 的基因组测序、组装与注释。研究表明,褐藻具有27-33条染色体,且保持了较好的宏基因组协同。通过生物信息学和实验方法,作者识别到物种中的雌性(U)和雄性(V)性别决定区。研究发现,这些物种共享一个相同的祖先性别染色体,尽管经历了高度重排仍保持其稳定性。系统进化研究表明,U/V 性染色体的诞生发生在约 450-224 万年前,通过抑制重组并包含雄性决定基因 MIN。部分物种缺乏 U/V 性染色体,但仍保留 MIN 基因。进一步分析表明,SDR 的基因数量和大小在物种间差异显著,并与基因数量和重复内容密切相关。研究还发现,SDR 边界在物种间有所变化,并伴随结构重排。综合结果表明,褐藻的 U/V 性染色体通过在含 MIN 的区域内抑制重组而进化,进一步研究可能有助于确定双性在这些物种中的存在。
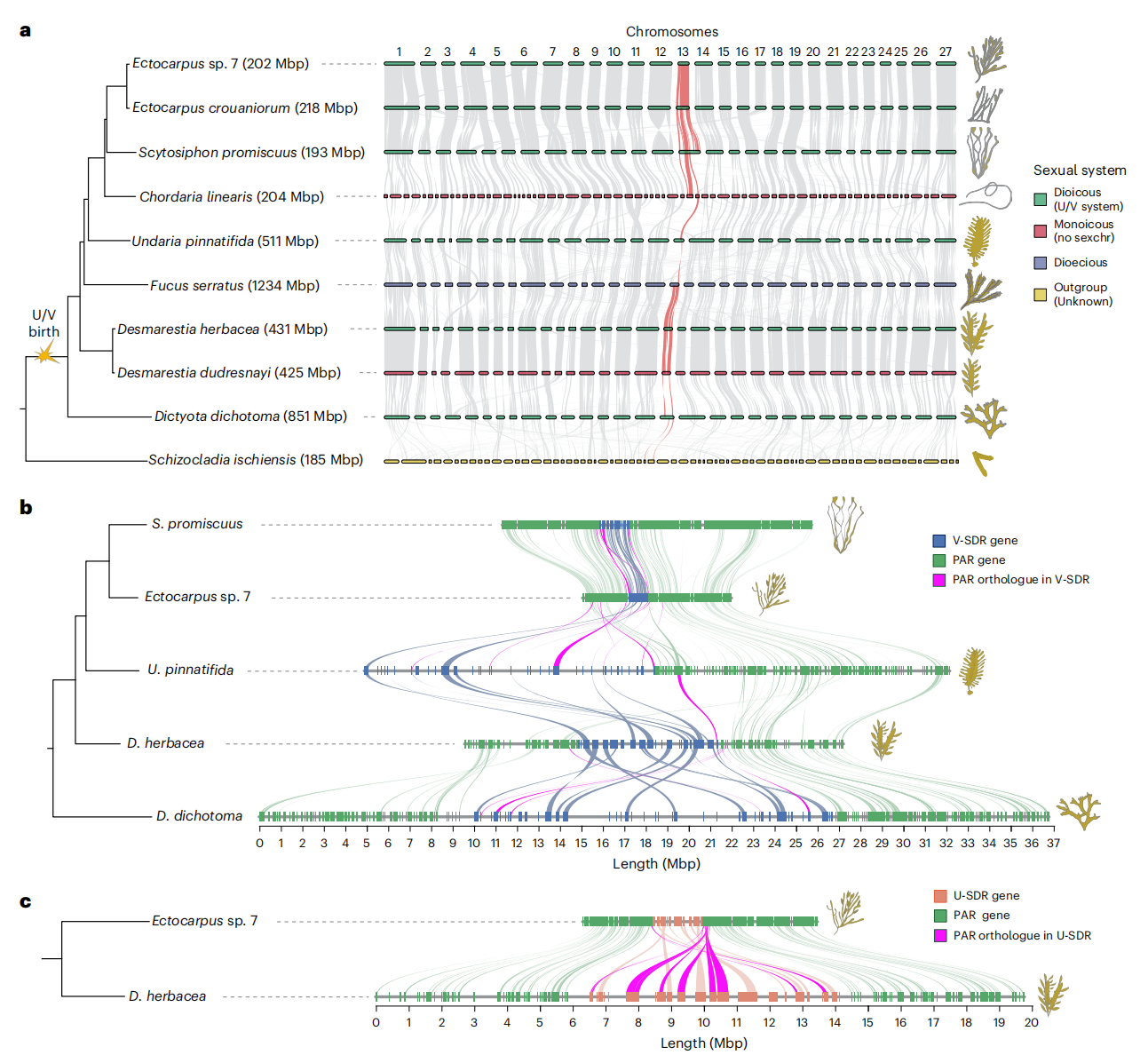
图1:褐藻U/V性染色体的起源
二、褐藻 U/V 性别决定区的进化:边界扩展与基因获得
研究发现,褐藻的U和V的SDR中含有同源基因,表这明它们源自共同的祖先区域。而在Ectocarpus sp. 7 和 D. herbacea 的 U/V-SDR 中,配子对和性别特异性基因的比率相似,尽管两物种的 U/V-SDR 基因总数不同。两物种的 SDR 区域中仅有十个基因为同源基因,其余则为物种特有基因。在 Ectocarpus sp. 7 中保留的一个基因在 D. herbacea 中已经丧失。两物种共有五对配子在雌雄染色体上都保留了副本,而另有三对配子在 D. herbacea 中仅保留了单副本,即雄性或雌性基因缺失。
不同于动物和植物,在褐藻的 U/V 系统中,性别决定区(SDRs)缺乏重组,加之基因移动和染色体重排破坏了共线性,因此进化层次难以识别。在 Ectocarpus sp. 7 与 D. herbacea 中,SDR 均发生倒位,可能是导致重组抑制的关键因素。对配子对同义替代率(Kₛ)的分析表明突变已接近饱和,限制了层次推断。但共享配子 Kₛ 值更高,显示其分化更早,而特有配子则较低。值得注意的是,D. herbacea 的 U-SDR 通过倒位扩展边界,吞噬了 Ectocarpus sp. 7 的部分伪常染色体区(PAR)基因,其中 12 个被整合并保留为配子,这支持了 SDR 通过嵌套倒位扩展的模式。此外,早期分化的 D. dichotoma 拥有更高的 V-SDR 基因含量。祖先 SDR 重建结果表明,褐藻 V-SDR 的演化主要依赖谱系特异性基因获得,而非基因丧失,其机制包括边界扩展、常染色体转位和谱系特异性基因新生。
研究发现,祖先V-SDR区域中包含七个基因,其中包括雄性决定基因 MIN 和六个与 U 染色体上的基因同源的基因。这些基因在 Ectocarpus sp. 7 中表现为配子对,但在 D. herbacea 中部分基因丧失,尤其是酪蛋白激酶和膜受体基因,这可能与性别系统丧失有关。尽管不同褐藻物种间的 U/V-SDR 基因数目不同,但所有物种的U和V SDR 中配子对及性别特异性基因的比例相对一致,这表明性染色体在各谱系中平行进化。研究还发现,V-SDR 的大小与性别二态性水平相关,性别二态性较低的物种保留了祖先的 V-SDR 基因,且基因变化较少。总体而言,褐藻的 U/V-SDR 主要通过谱系特异性基因获得而非基因丧失,基因获得与性别二态性发展密切相关。
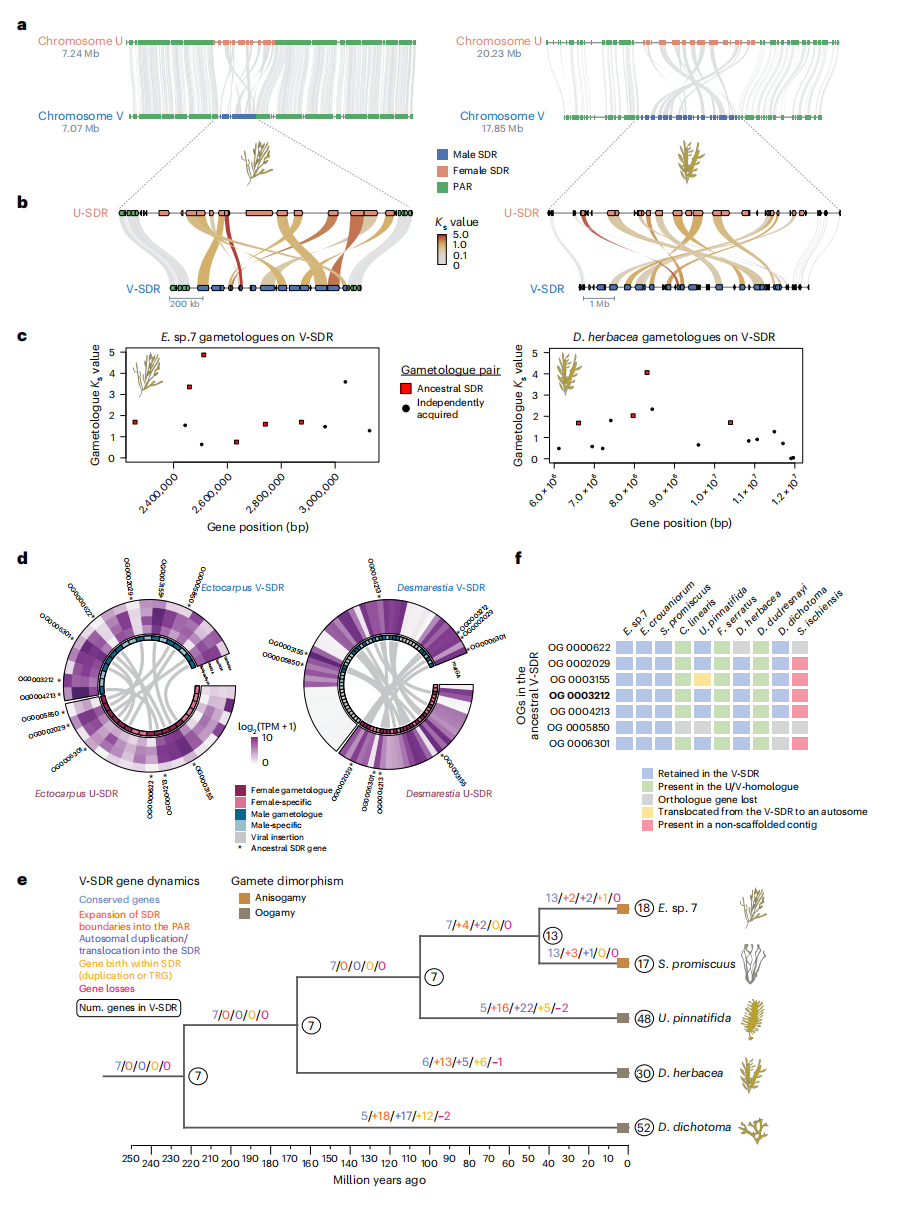
图2:来自祖先SDR的特定谱系U/V-SDR扩展及其与性别二态性的关联
三、褐藻U/V性染色体的结构特征和进化动力学
作者研究了褐藻 U/V 性别染色体(V-SDR 和 PARs)的结构特征及其进化动态。研究发现,V-SDR 区域重复序列丰富,但基因较为稀缺,而 PARs 区域也富含重复元素,尤其是在基因组扩展的物种中。这些区域的基因同源性低于常染色体,可能与分类限制基因(TRGs)数量的增加有关。TRGs 是指仅在某些物种或物种群体中存在的基因,反映了性别染色体上的基因多样性。通过系统发育分析,作者发现所有双性物种的性别染色体都富含 TRGs,且在 V-SDR 较大的物种中,TRGs 的富集更为显著。
进一步分析显示,性别染色体上的 TRGs 比物种共同祖先的 TRGs 更年轻,这表明 TRG 的富集是在不同物种中独立演化的结果。整体来看,这些发现揭示了性别二态性与基因进化之间的紧密关系,为褐藻 U/V 性别染色体的复杂演化过程提供了深刻的见解。
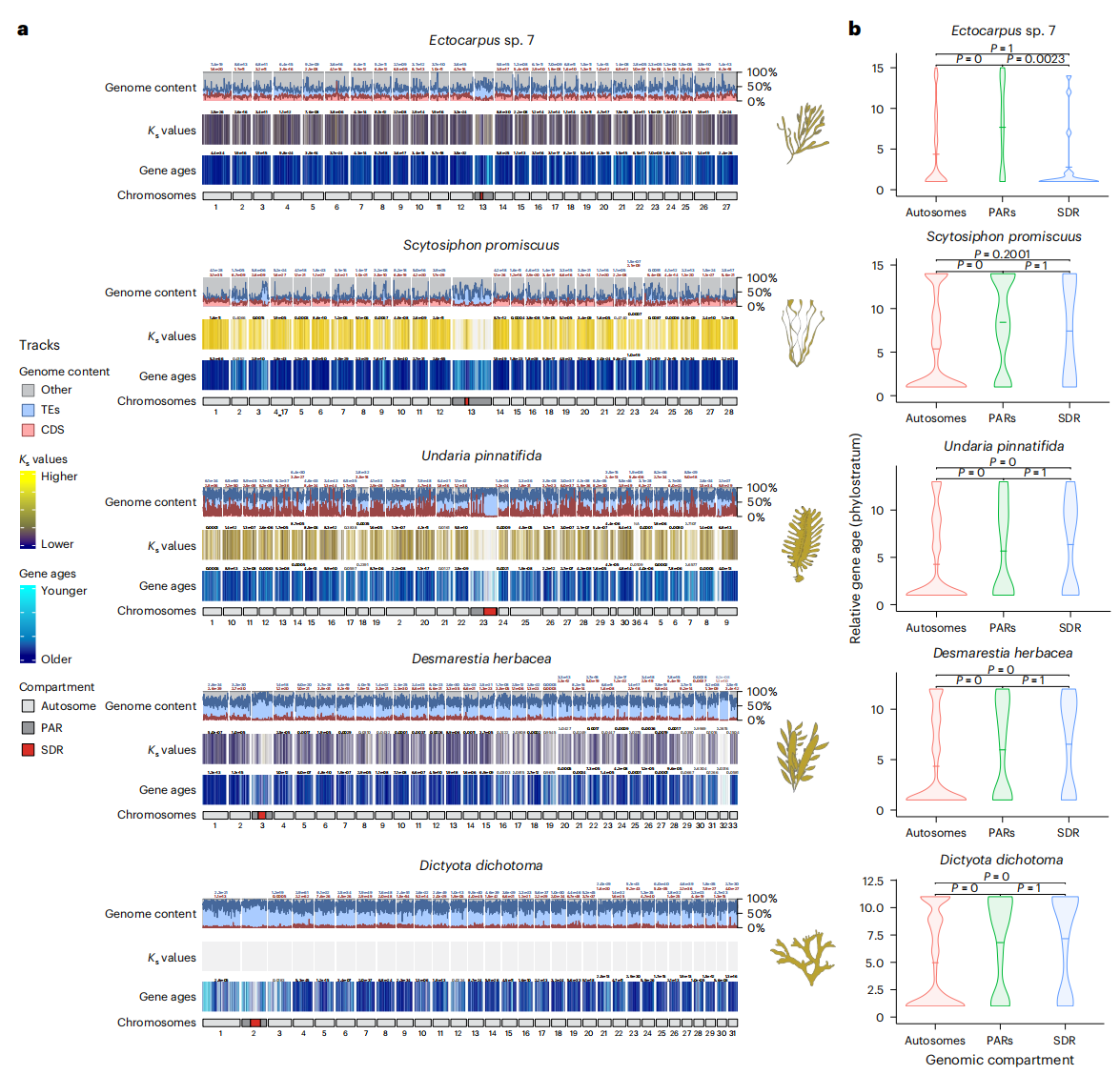
图3:U/V 性染色体富含褐藻特有基因
四、雌雄体丧失后U/V性染色体的命运
作者研究发现,褐藻U/V性别决定系统的演化呈现出两种典型模式。在C. linearis和D. dudresnayi这两个独立进化的物种中,U/V系统丢失后演变为雌雄同体,其"前"性别染色体表现出明显的雄性起源特征:保留了包括MIN基因在内的大量V-SDR基因,但丢失了多数U-SDR基因,同时伴随显著的结构重排。值得注意的是,这些V-SDR基因在生殖期仍保持活跃表达,表明其在性别分化中的功能得以保留。更为特殊的是F. serratus的案例,作者发现该物种经历了从单倍体到二倍体性别决定的罕见转变。虽然其"前"性别染色体已丧失典型U/V特征,但MIN等五个V-SDR基因仍维持严格的雄性特异性表达模式。这一发现提示,古老的V-SDR基因可能被重新整合至新的二倍体性别决定通路中。
通过比较研究,作者揭示了一个重要规律:尽管原始U/V系统可能丢失,但其核心基因(如MIN)表现出显著的功能保守性,只是通过不同的染色体结构和调控机制继续发挥作用。这一发现为理解性别决定系统的演化提供了新的见解。
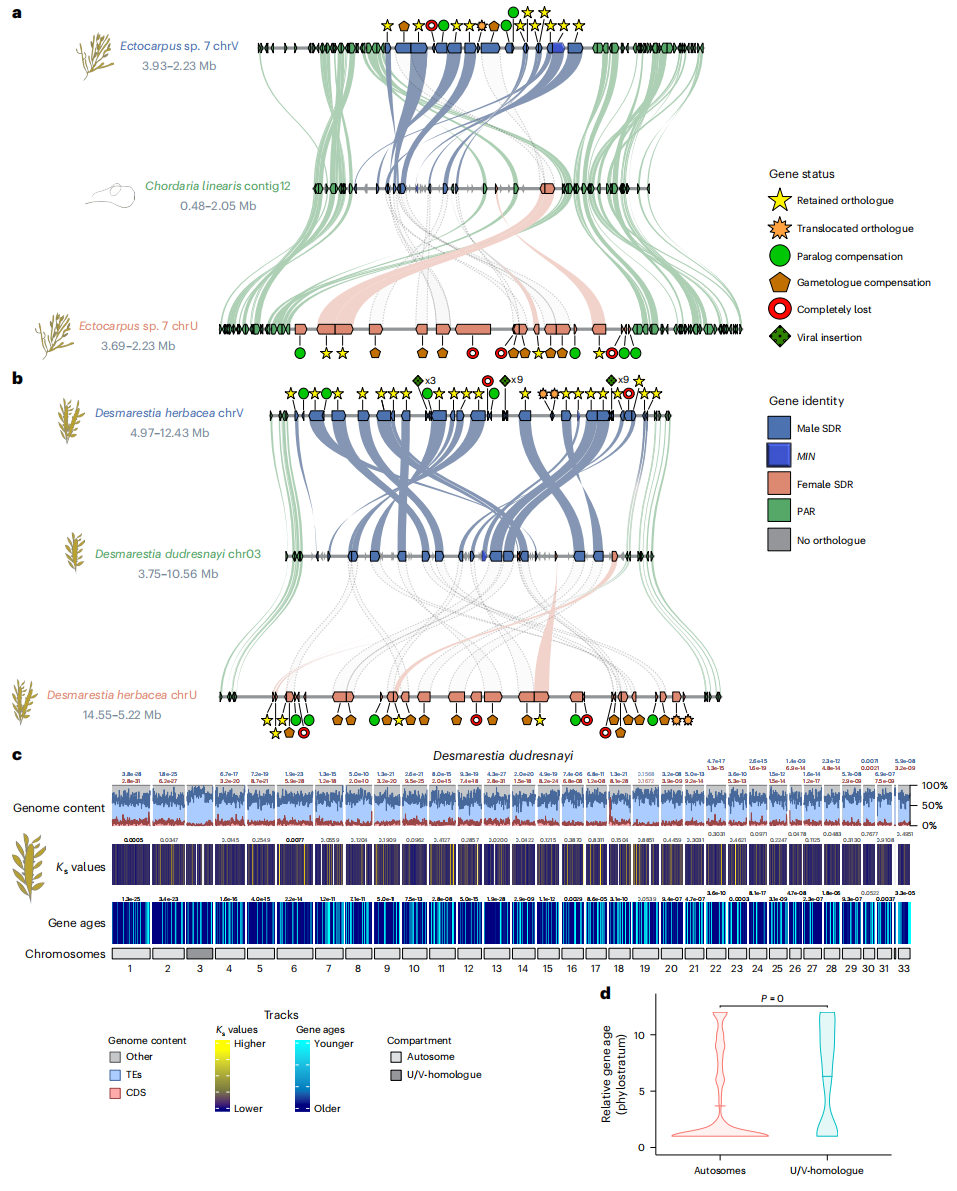
图4:性染色体从雌雄株向雌雄同体转变过程中的演变
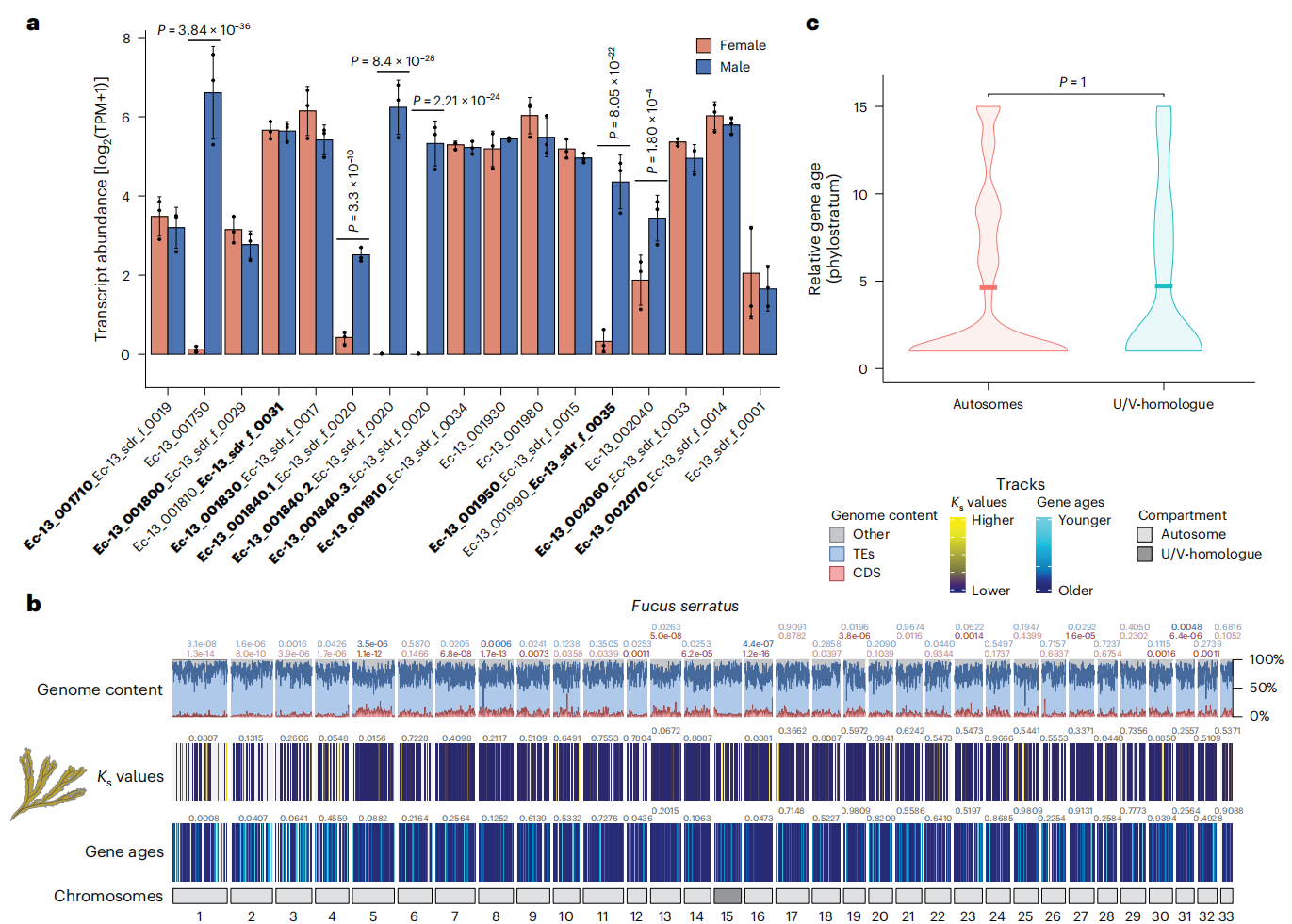
图5:从单倍体到二倍体性别决定的转变
结语
本研究重点分析了9种褐藻的基因组,并追溯了U/V性染色体的演化历史。进化分析发现,性别决定区域起源于4.5-2.24亿年前,由包含雄性决定基因MIN的区域重组抑制形成。尽管7个祖先基因高度保守,但嵌套倒位在不同谱系中独立扩展了性别位点。研究发现分类限制基因在U/V染色体中频繁演化,并揭示两种U/V区域变异模式:单性物种通过祖先雄性获得U特异性基因实现趋同进化;Fucus dioecious雌雄异株系统则依赖新的性别决定基因调控V特异性基因。这两种情况导致了U和V染色体的消亡,以及它们特有基因组特征的侵蚀。
华命生物产品服务一览

华命生物目前已开通微信公众号、抖音、知乎、B站、小红书等线上平台,欢迎感兴趣的老师扫码关注了解更多内容!
