

MP精选 | 被子植物着丝粒演化探究
着丝粒是维持染色体正确分离与基因组稳定的关键结构,但其序列与结构在物种间却高度多样、演化迅速,形成“功能保守而序列快速演化”的悖论。由于着丝粒DNA由大尺度卫星重复与转座元件交错构成,长期难以被常规组装解析,而T2T测序的进展显著推动了着丝粒研究。如今,着丝粒变异被认为可驱动基因组创新与生殖隔离,参与核型演化与物种形成。被子植物因基因组大小及重复序列含量差异巨大,是研究该问题的理想体系。
华中农业大学研究团队于2025年12月18日于国际著名期刊Molecular Plant上发表了标题为“Evolutionary innovations and genetic diversity in angiosperm centromeres”的研究性论文,通过整合多种被子植物的高质量着丝粒、泛基因组和群体数据,揭示了在功能高度保守的背景下,卫星重复序列与转座元件的协同演化驱动着丝粒快速分化的过程。

一、被子植物着丝粒组成与结构的演化多样化
为解析被子植物着丝粒的演化多样化,研究分析了17种二倍体物种、400余个近乎无缺口的着丝粒,发现其大小具有高度可塑性,物种间差异显著:拟南芥着丝粒较为保守,而马铃薯则高度变异。着丝粒大小与基因组大小和染色体长度呈弱正相关,但以 CENH3 占据范围估算时相关性更强,且存在小基因组物种着丝粒反而较大的例外。组成上,不同物种间卫星 DNA 与转座元件比例差异巨大,着丝粒结构呈现由“卫星主导的简单型”向“TE 富集的复杂型”转变。结构分析进一步表明,复杂着丝粒通常具有更大的 CENH3 富集域,并在卫星与 CENH3 分布上表现出多样组合,反映着丝粒在谱系特异演化压力下于序列结构约束与表观遗传功能之间实现动态平衡。
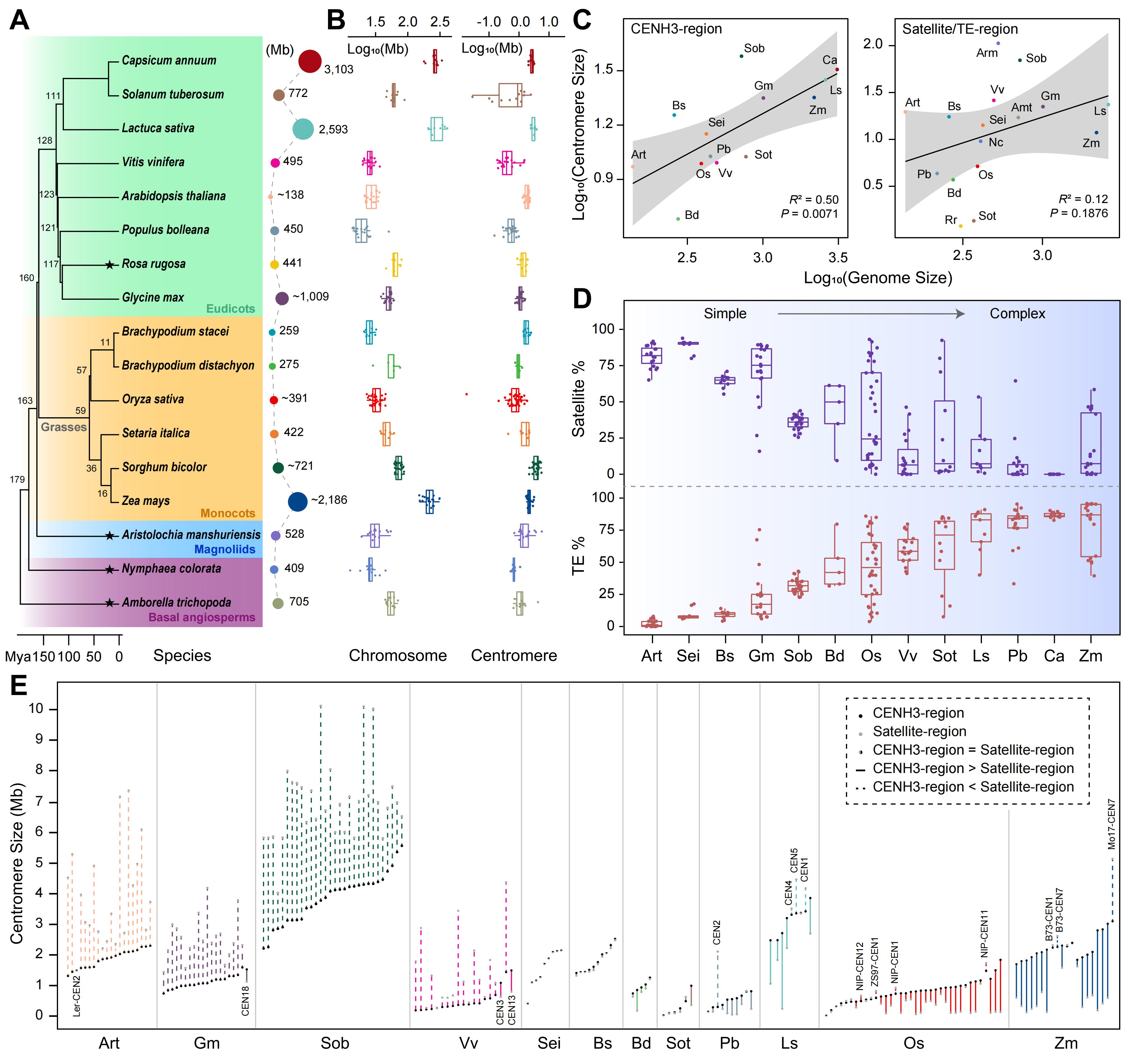
图1. 不同被子植物基因组中着丝粒景观的综合分析
二、被子植物着丝粒卫星阵列的差异化演化轨迹
研究系统比较了多种被子植物中着丝粒卫星重复序列的组成、结构及其演化特征,结果表明不同物种的着丝粒通常由彼此无关的卫星重复家族构成。尽管多数物种的卫星单体长度集中在100-200 bp、符合CENH3核小体结合的结构需求,但仍存在单体长度显著延长或多样化的特例,显示着丝粒结构具有较高的可塑性。进一步分析发现,倾向于简单型着丝粒的物种通常具有更高丰度、序列更均一的卫星重复,易在染色体内发生同质化并形成较长的高阶重复结构;而复杂型着丝粒中的卫星重复则数量较少、结构更为碎片化。与此同时,卫星序列的均一性与CENH3的富集程度呈正相关,说明卫星重复的结构稳定性可能有助于维持着丝粒的功能。
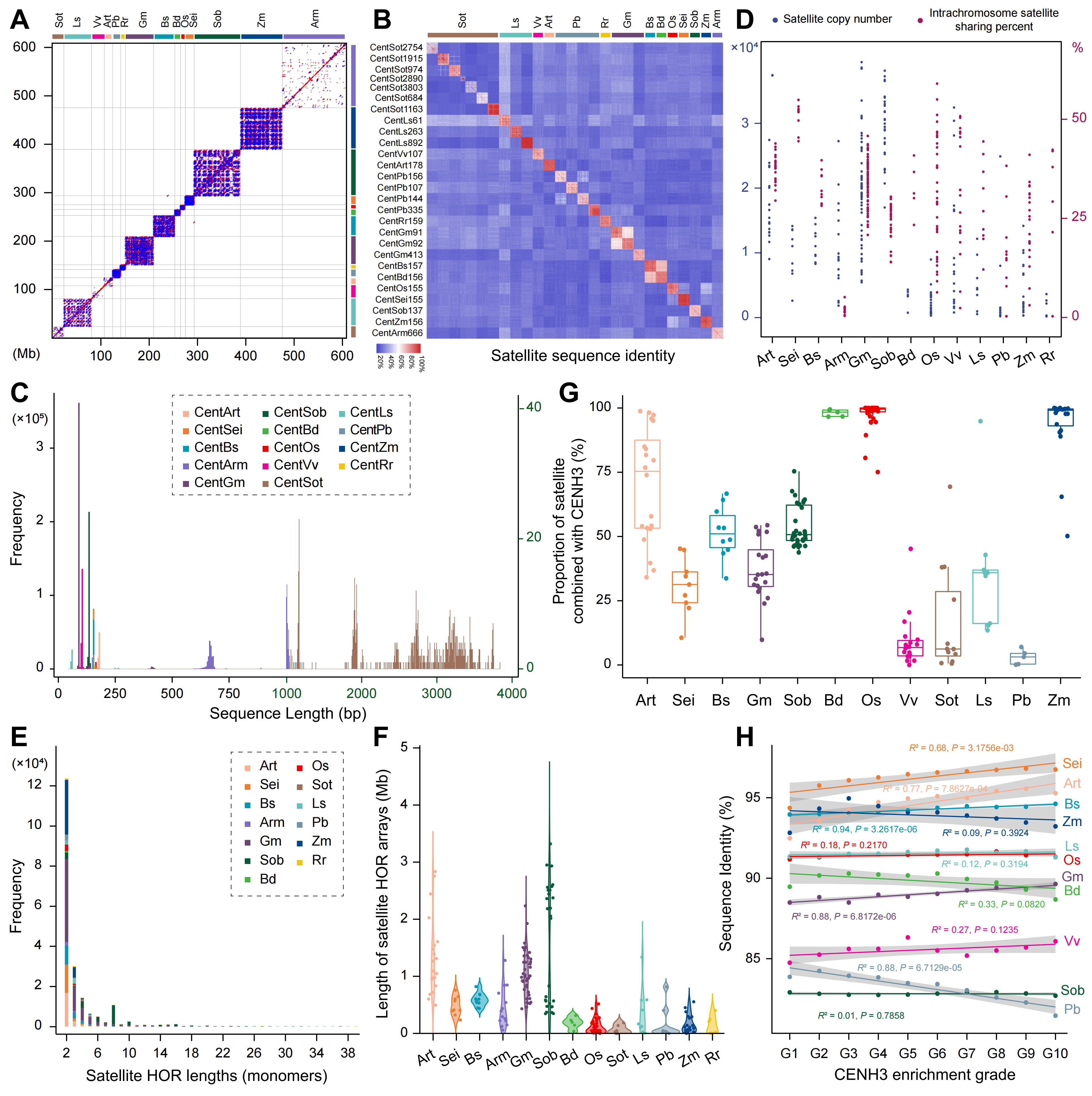
图2. 不同被子植物基因组中着丝粒卫星序列的精细结构
三、转座子动态驱动被子植物间着丝粒结构变异
为了进一步阐明被子植物着丝粒结构多样化的机制,作者进一步系统比较了多种物种中着丝粒相关转座子(TE)的组成、演化动态及其与CENH3的关系。结果显示,不同物种着丝粒中转座子的丰度和类型差异显著,复杂型着丝粒普遍富含转座子并具有较高比例的TE–CENH3结合,而简单型着丝粒则以卫星DNA为主要结构成分。多数被子植物着丝粒由LTR反转座子,尤其是Gypsy超家族成员占据主导地位,这些转座子在不同谱系中经历了相互独立的扩增与分化过程。进一步分析表明,着丝粒区域显著富集较新的转座子插入,但不同物种在转座子更新模式上存在明显差异,表现为持续扩增或长期保留的不同策略。转座子插入对着丝粒结构具有显著影响,高转座子密度往往伴随卫星序列均一性的降低,并改变CENH3核小体的排列方式。在转座子富集型着丝粒中,部分转座子元件可作为CENH3装载的结构支架。
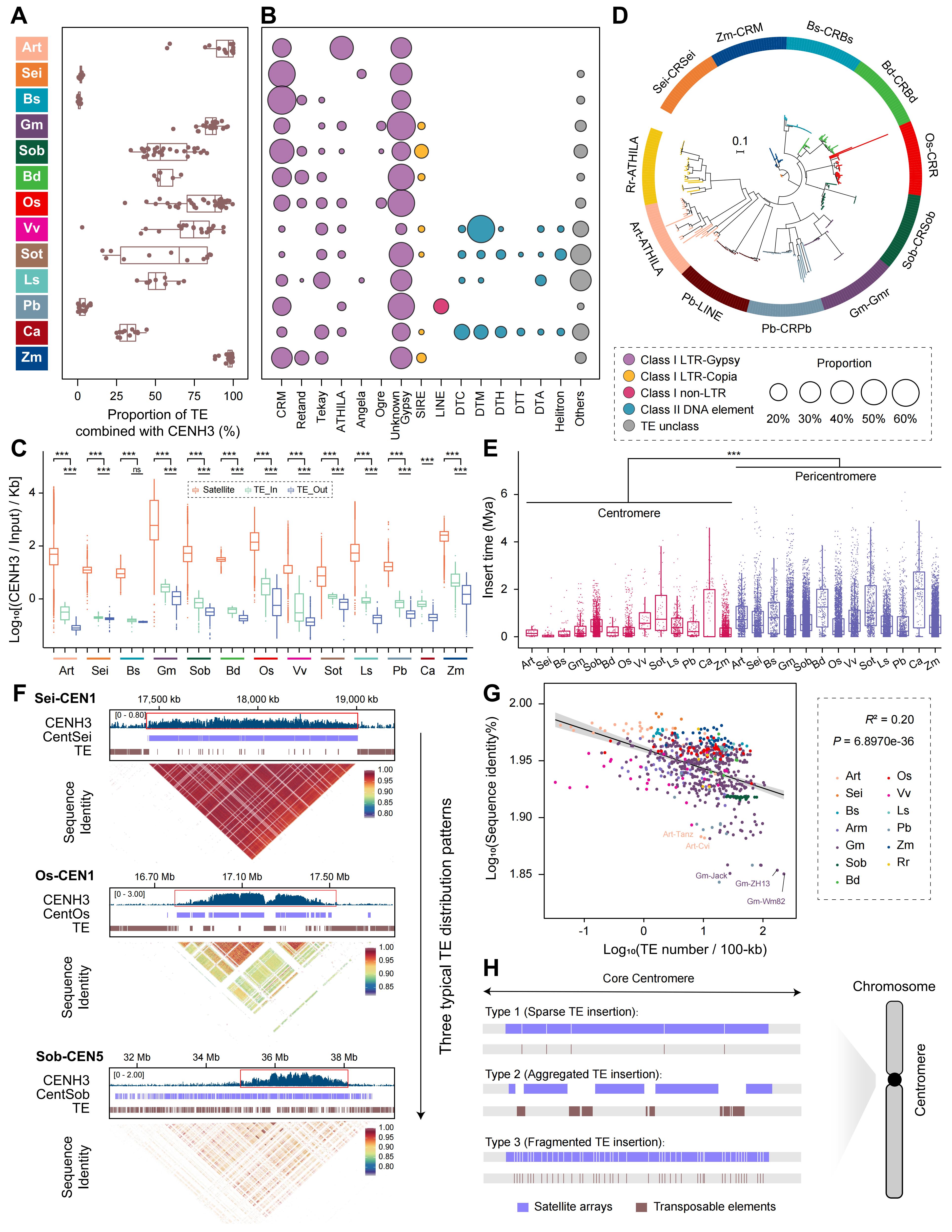
图3. 转座子入侵格局及其与被子植物着丝粒结构多样性的关联
四、被子植物着丝粒的群体水平多样性分析
群体水平的比较分析显示,被子植物种内着丝粒在大小、结构组织和序列组成上普遍存在显著多态性,但不同物种的分化程度存在明显差异。葡萄和玉米在不同品系间表现出强烈的着丝粒结构分化,而高粱的着丝粒在序列和结构层面高度一致,显示出较强的稳定性。进一步分析表明,种内着丝粒结构变异主要来源于卫星重复和转座子驱动的动态扩增与收缩,并伴随卫星方向重排及高阶重复结构重塑。尽管着丝粒整体具有较高的结构流动性,部分染色体的着丝粒在不同材料中仍保持高度保守,表明其受到功能性约束。群体层面的序列比较显示,着丝粒区域的序列一致性普遍低于周着丝粒区域,反映其较快的演化速率。着丝粒重新定位在部分物种中频繁发生,尤其在经历过多倍化的物种中更为显著,说明转座子活性、多倍化历史及卫星重复动态共同塑造了着丝粒在群体中的结构稳定性与空间位置。
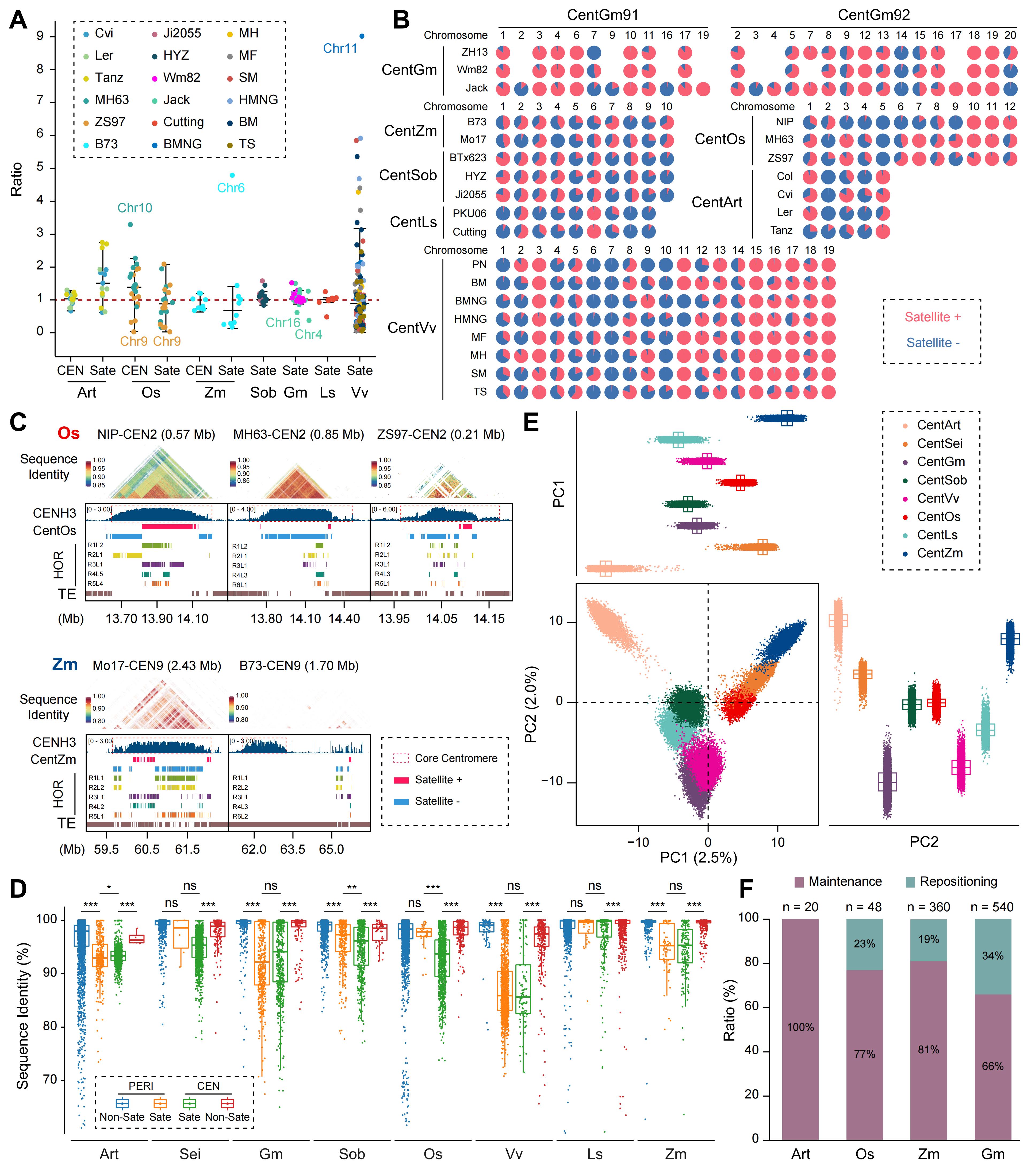
图4. 不同被子植物基因组中群体水平的着丝粒遗传和表观遗传多样性
五、被子植物着丝粒在物种形成过程中的渐进式分化
通过整合拟南芥、水稻、短柄草和玉米等属的284个基因组数据,研究发现着丝粒卫星重复的分化程度与物种分化时间呈现清晰关联:在近期分化的同属物种中(<5 Mya),着丝粒卫星整体保持高度保守,不同物种通常共享相同或高度相似的重复家族,仅存在有限的序列多态性和扩增差异;随着分化时间延长(约5-10 Mya),共享卫星重复逐步分化为与物种边界一致的系统发育亚分支,形成明确的物种特异性着丝粒谱系;在更深层次分化的类群中(>10 Mya),着丝粒组成经历显著重构,表现为原有卫星重复的丢失或被全新的重复家族所替代。进一步分析显示,这一分化过程常与染色体融合及染色体数目变化等核型重排事件相伴发生,并可能加速着丝粒序列的更新。此外,多倍化在保留祖先着丝粒特征的同时,引入新的变异并促进进一步分化。总体来看,着丝粒序列在不同演化时间尺度上持续更新,其渐进式分化与物种形成过程高度同步,使其成为解析被子植物系统发育关系与宏观演化历史的重要基因组标记。
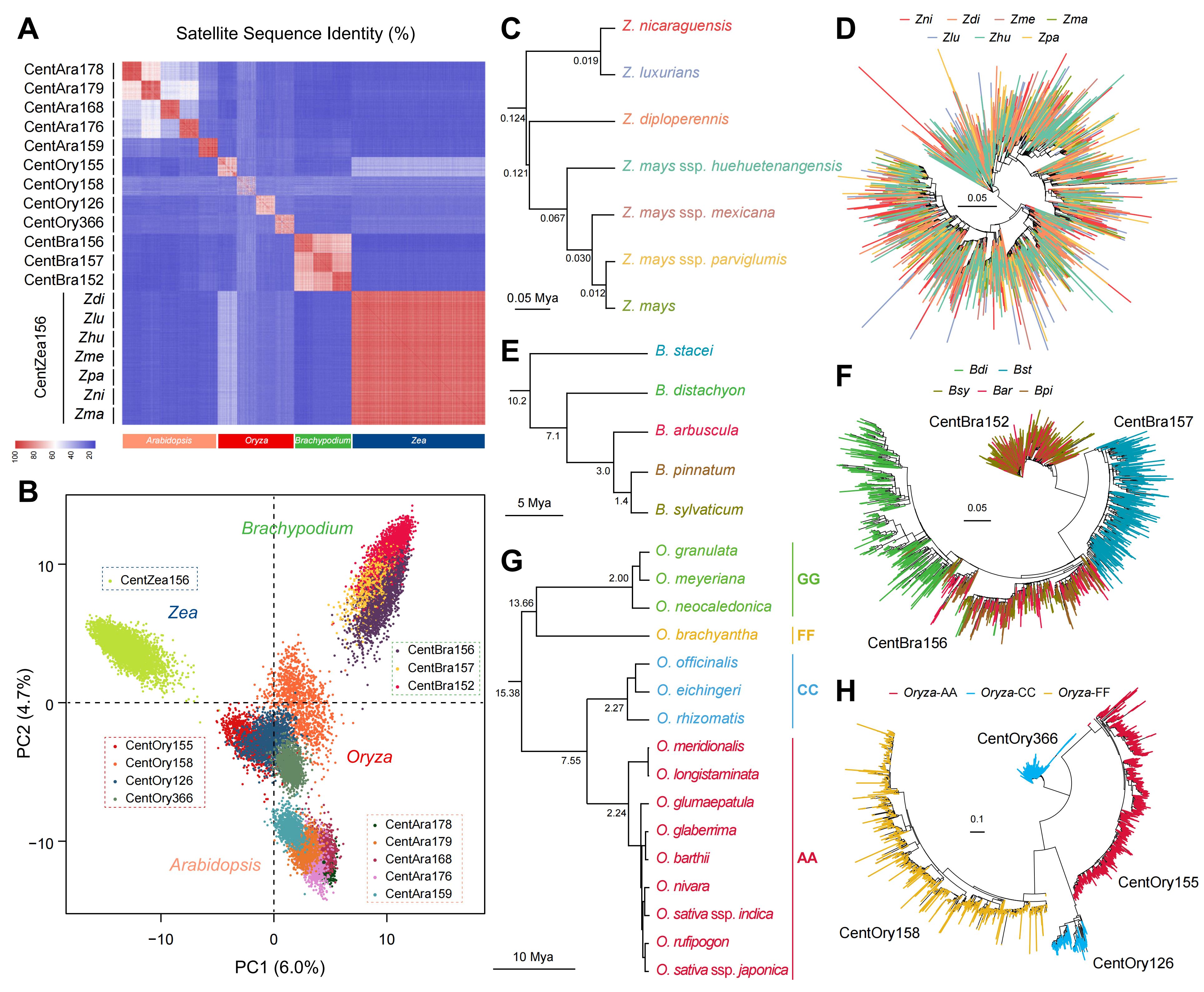
图5. 拟南芥属、短柄草属、水稻属和玉米属着丝粒卫星重复序列的起源与演化
结语
本研究系统分析了被子植物着丝粒在演化过程中的结构创新与多样性。基于17种被子植物、400余个高质量着丝粒及泛基因组和群体数据,作者揭示了着丝粒在功能保守背景下仍具有快速序列演化特征。结果表明,谱系特异性的卫星重复与转座元件共同塑造了着丝粒结构,其中转座元件的插入与扩增推动了结构复杂化、序列更新及位置重排。着丝粒在种内高度可塑,在种间经历由渐进分化到完全替换的连续演化,并常伴随染色体结构重排,表明其演化受基因组动态、染色体重塑与选择压力的协同影响,并可能参与核型演化与物种形成。