

PBJ精选 | 龙眼near-T2T基因组解码反季开花现象
龙眼(Dimocarpus longan Lour.)是重要的热带亚热带果树,具有显著的经济和药用价值。氯酸钾(KClO₃)诱导的反季节开花技术显著提高了龙眼周年生产能力,但其分子调控机制尚不清楚。受限于童期长和育种周期久,龙眼遗传改良进展缓慢,目前基因组资源仍然匮乏,主栽品种石硖虽对氯酸钾诱导开花高度敏感,但其基因组尚未系统解析。
福建农林大学研究团队于2025年11月4日在Plant Biotechnology Journal上发布标题为“Improved Longan Genome Assembly Reveals Insights Into Flowering Mechanisms”的研究性论文,本研究构建了石硖龙眼高质量near-T2T基因组,揭示了氯酸钾诱导的反季节开花主要通过抑制 DlDDF1 并激活赤霉素信号通路实现,而自然开花则主要依赖季节性环境信号调控的生理机制。

一、near-T2T基因组组装与注释
本研究估算石硖龙眼基因组大小为441.5Mb,基于PacBio HiFi(51×)和Hi-C(200×)测序数据,使用HiFiasm完成基因组组装,获得98个contig,contig N50为28.3Mb,总长度441.5 Mb,其中99.2%序列被锚定至15条假染色体。组装基因组BUSCO完整度为98.4%,QV达68.1。共注释得到29,325个蛋白编码基因,BUSCO完整度为97.7%,预测到26个端粒和15个着丝粒,仅3条染色体上存在少量gap。非编码RNA注释获得137个miRNA、885个snRNA、437个tRNA和1,627个rRNA,96.4%的基因实现功能注释。转座元件占基因组的55.0%,以反转座子为主。
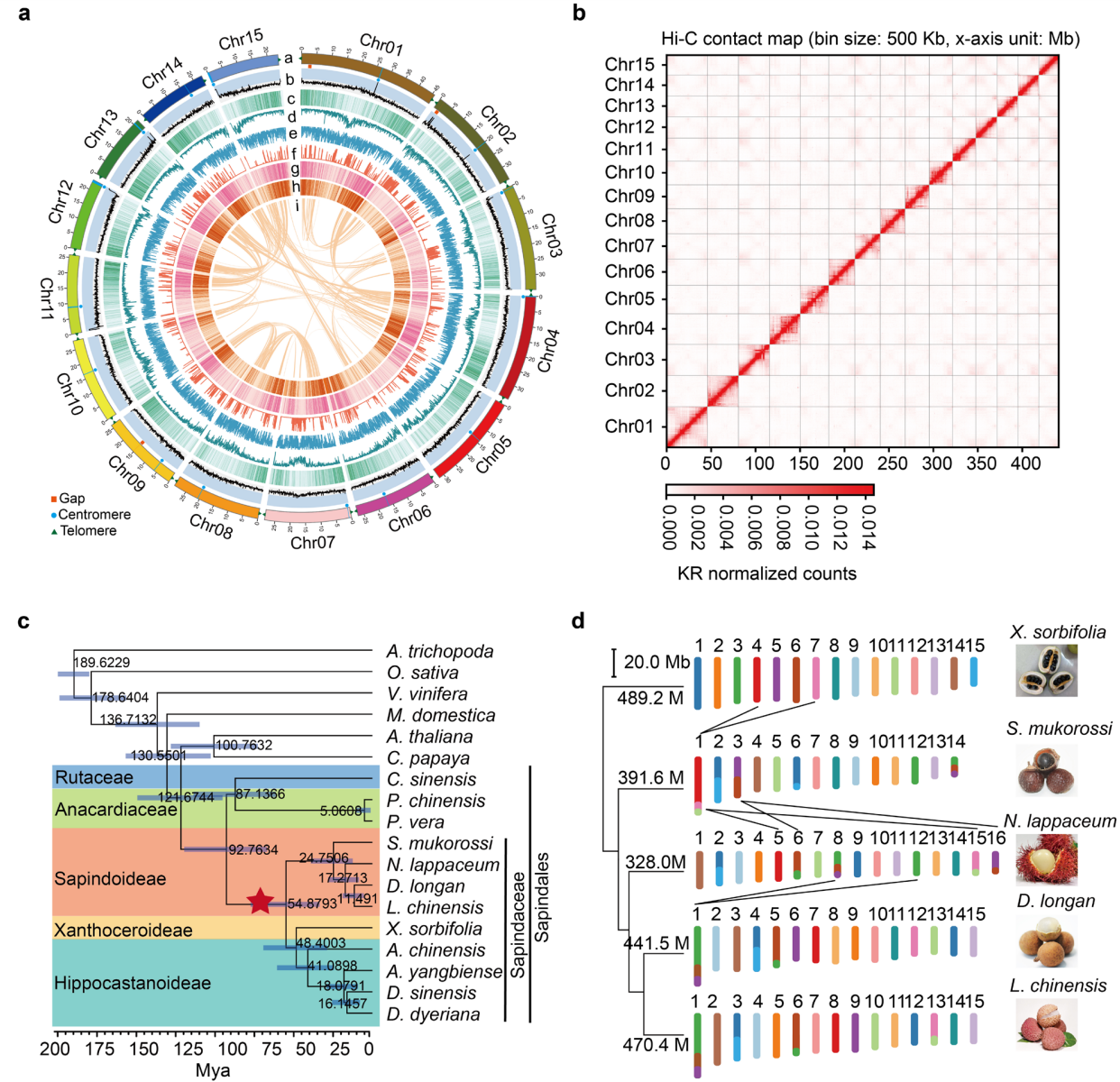
图1:石硖龙眼near-T2T基因组特征、Hi-C图谱及核型进化
二、比较基因组分析
系统发育与共线性分析表明,龙眼与荔枝、无患子、文冠果和红毛丹亲缘关系密切,其染色体数目受融合与断裂事件影响,在 14-16条之间发生变化。尽管各物种非重复序列基因组大小相近,但总基因组大小差异主要由重复序列含量不同所致,龙眼基因组中以 Gypsy 类转座元件占优势。比较基因组分析共鉴定出 486 个龙眼特异性基因家族和 3206 个特异基因,显著富集于病原响应、糖代谢及黄酮/黄酮醇生物合成通路,且相关基因在花芽中高表达。与荔枝相比,龙眼基因组中存在大量倒位、重复和易位等结构变异,这些区域富集植物抗病和黄酮合成相关基因,并包含多个花期调控基因(如 DlFT1、DlELF4 和 DlSVP),表明结构变异及转座元件扩增可能参与龙眼花期调控与性状分化。
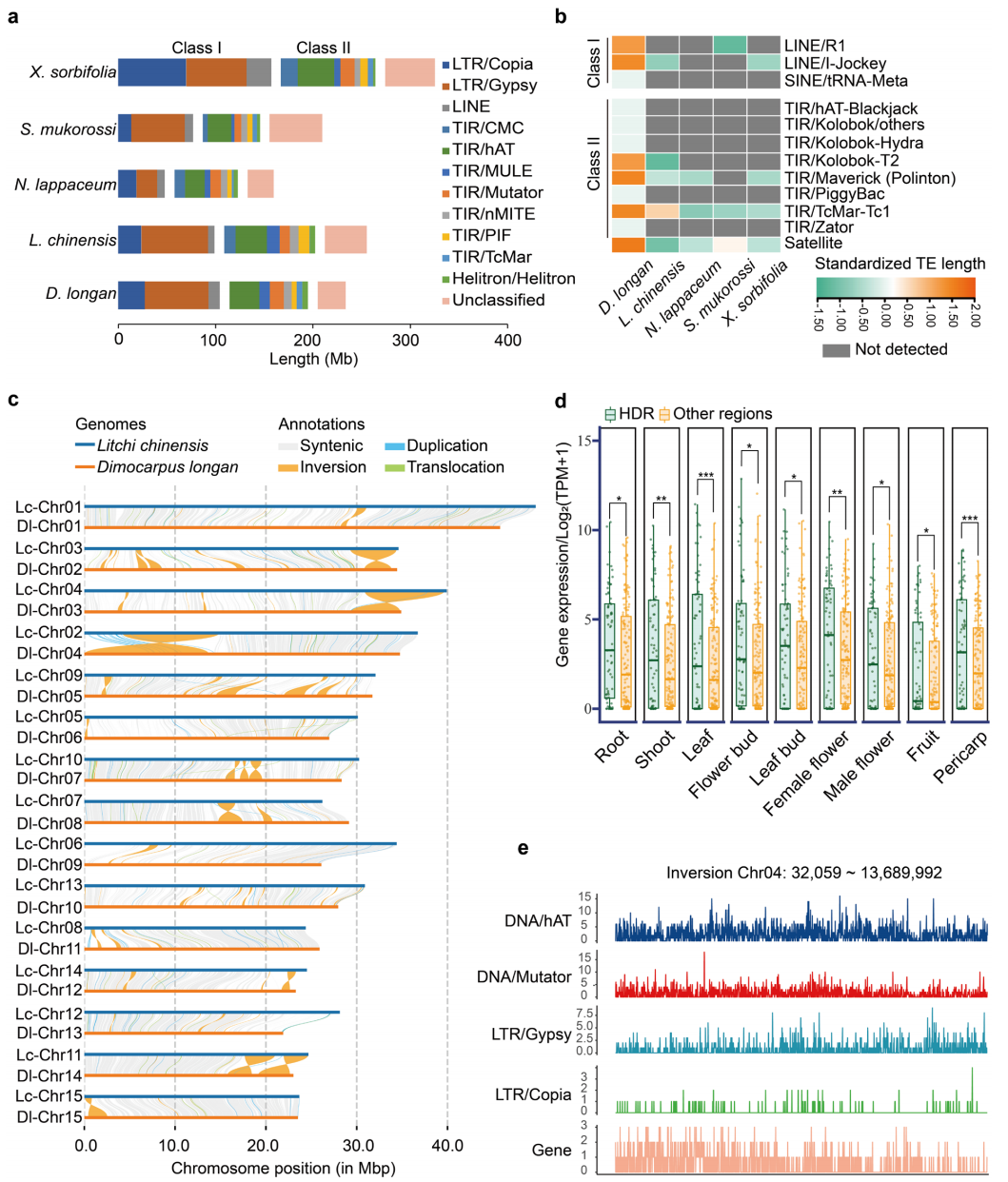
图2:龙眼和荔枝的可转座元件及结构变异比较基因组分析
三、龙眼的自然花芽转变
在自然当季条件下,对5个龙眼品种的花芽和叶芽进行转录组分析,共鉴定到13673个差异表达基因,其中6522个在花芽中上调、5922个下调,并分别有568个上调基因和919个下调基因在5个品种中共享。WGCNA分析显示,上调基因构成高置信度共表达网络,枢纽基因主要涉及光周期和自主开花途径;下调基因则多与胁迫响应和花发育负调控相关。基因调控网络分析表明,自然花芽转变涉及光周期、自主和春化途径,并与赤霉素及糖信号通路互作,其中DlSVP和DlTFL1等抑制因子受到抑制,而DlFT1、DlAP1和DlLFY等促花基因显著上调。常年开花品种“四季蜜”中,昼夜节律和光周期相关基因持续高表达,提示光周期调控网络的持续激活可能是其常年开花的重要分子基础。
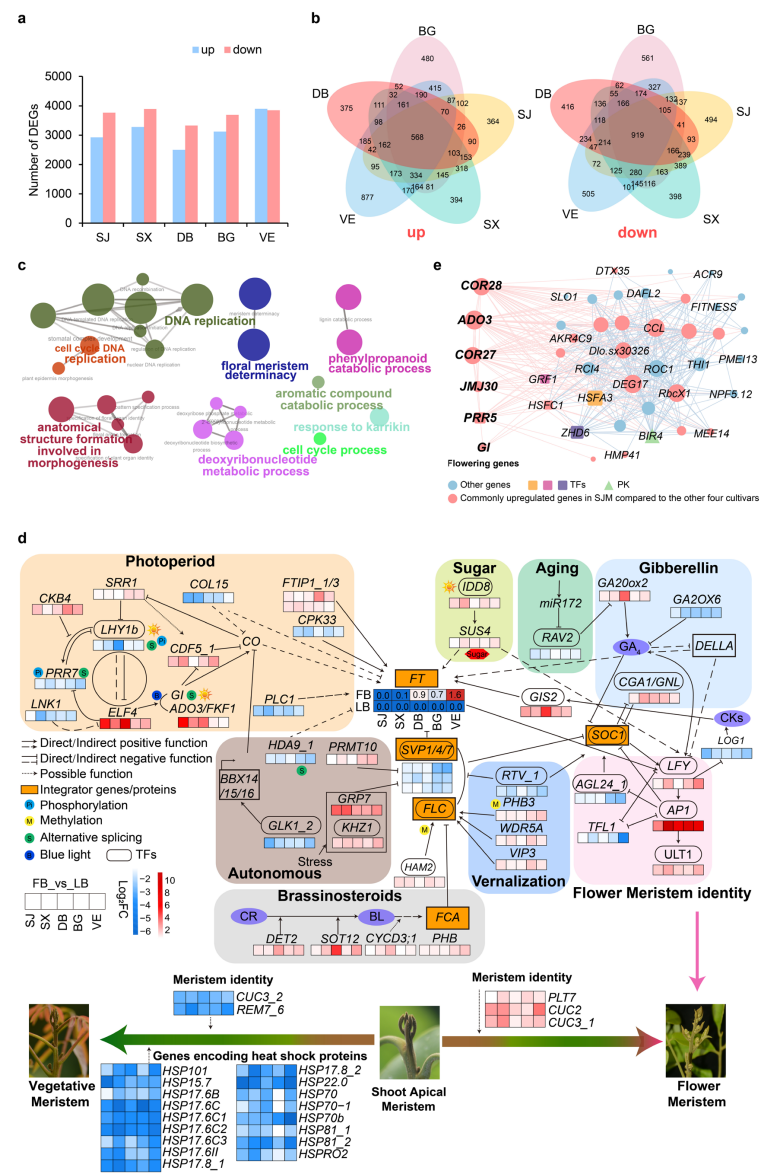
图3:龙眼自然花期转变的转录组动态
四、氯酸钾诱导龙眼反节开花的转录调控机制
在石硖龙眼中施用氯酸钾(KClO₃)后,花芽诱导率在处理后41天(DAT)和54DAT分别达到64%和85%,显著高于对照组。对处理组和对照组花芽在10个时间点进行转录组分析,共鉴定到2,851个差异表达基因,包括193个转录因子和155个激素相关基因。多数ERF、WRKY、MYB等转录因子在早期下调,而MIKC-MADS家族成员在后期上调。基因调控网络分析显示,赤霉素(GA)通路在诱导初期即被激活,其中GA生物合成抑制因子DlDDF1在5DAT显著下调,而DlGA3ox2、DlWRKY75_2及抑制GA₄降解的DlIAA7s上调,同时GA失活基因DlGA2ox4/6在后期被抑制。10-41DAT期间,自主途径基因上调,而DlSVPs和DlFLCs等花期抑制因子受到抑制,随后DlFT1、DlAP1、DlLFY等花分生组织和花器官身份基因依次激活。WGCNA分析进一步鉴定出与早期诱导、中期花起始及后期花器官发育相关的共表达模块,表明KClO₃诱导开花是由GA信号、胁迫响应及自主途径协同调控的过程。
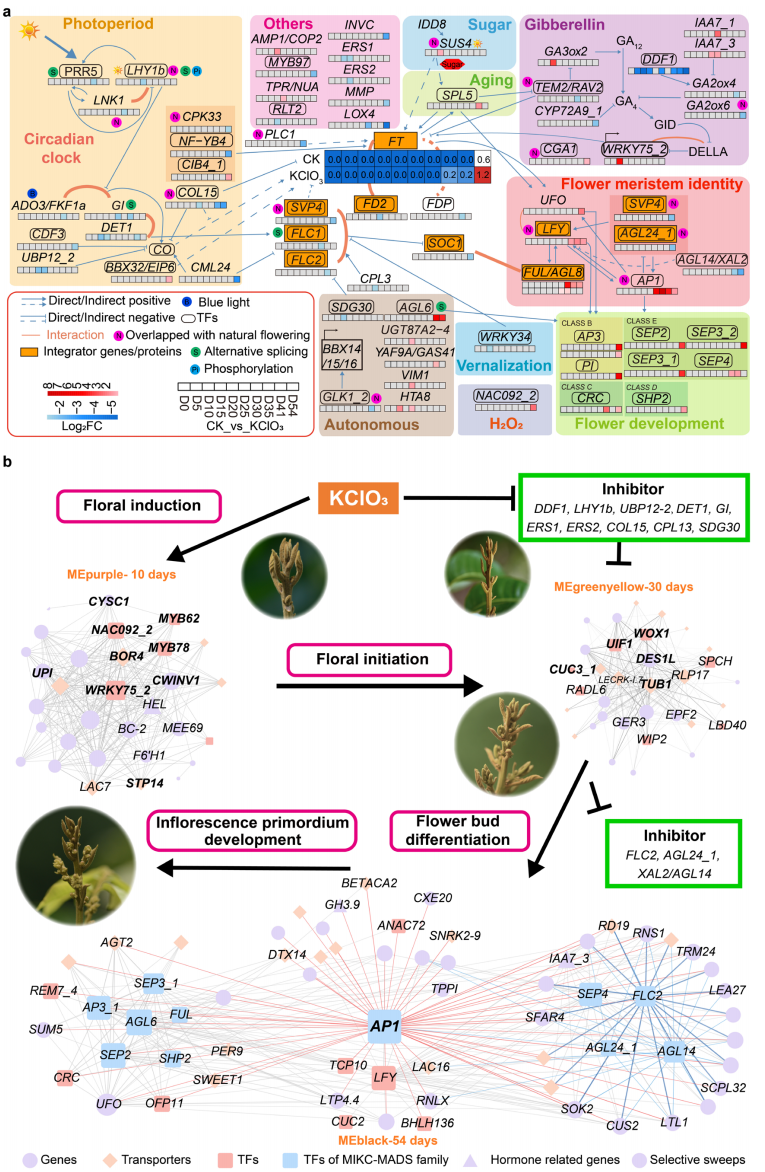
图4:KClO3诱导龙眼早花的转录组分析
五、DlDDF1 在开花调控中的功能验证
研究进一步系统验证了DIDDF1在开花调控中的功能,鉴于DIDDF1对KClO₃处理表现出最早响应,作者推测其参与KClO₃诱导的开花调控过程。通过构建DIDDF1过表达拟南芥材料发现,与野生型相比,转基因植株表现出显著的迟花表型,平均开花时间延迟约4天,并伴随莲座叶数增加及叶片形态异常,这表明DIDDF1在正常条件下对开花具有负调控作用。进一步的KClO₃处理结果显示,过表达植株的迟花表型可被部分恢复,且其对高浓度KClO₃所致生长抑制的敏感性明显低于野生型。激素分析表明,DIDDF1过表达显著降低GA₉ 及生物活性GA₄ 水平,而KClO₃处理可部分逆转该变化。转录水平分析进一步显示,KClO₃处理抑制DIDDF1表达的同时,显著上调AtFT、AtAP1和AtLFY等关键开花基因。综合来看,DIDDF1可能通过调控赤霉素代谢及开花通路基因表达,参与KClO₃介导的开花调控机制。
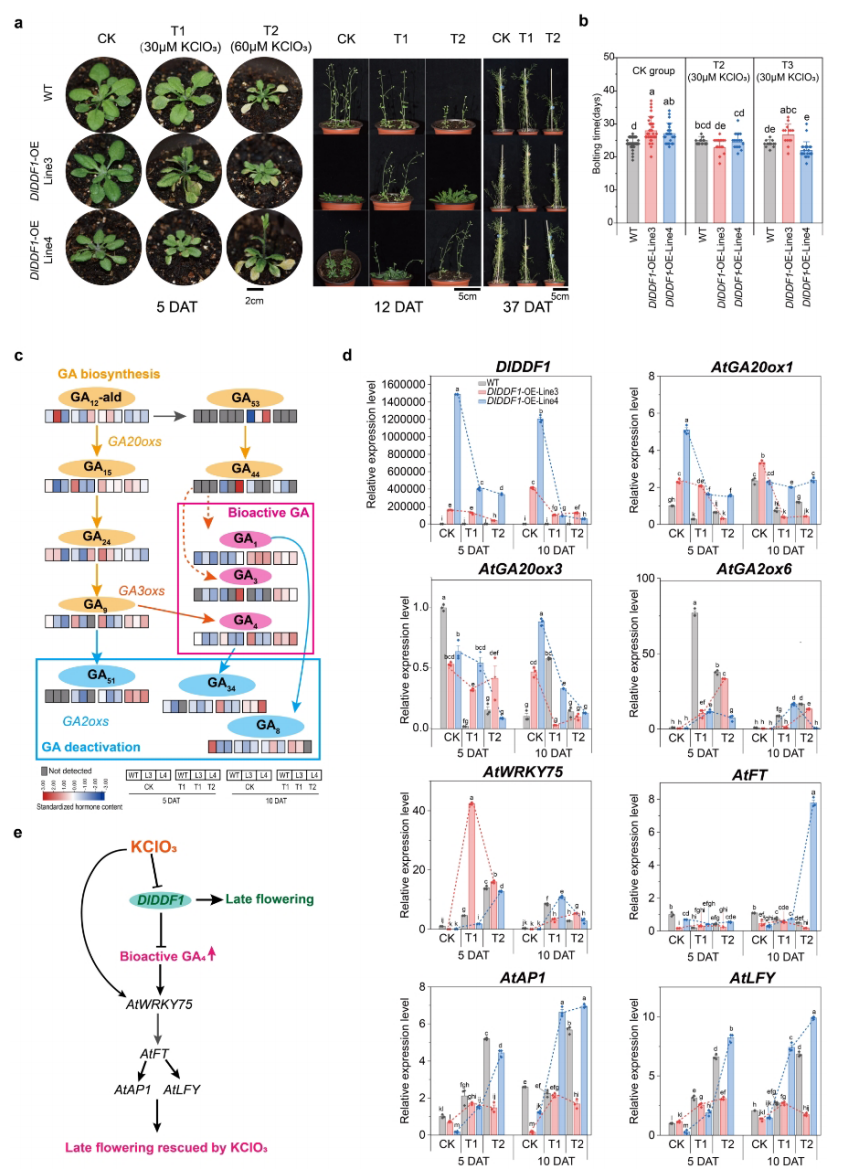
图5:DlDDF1过表达和KClO3处理对拟南芥的影响
结语
本研究构建了龙眼高质量near-T2T基因组,并结合比较基因组与多品种芽转录组分析,系统解析了龙眼自然成花与KClO₃诱导反季开花的分子调控机制。结果表明,自然成花主要受光周期、春化和自主通路调控,而KClO₃诱导开花则更早激活赤霉素信号并伴随氧化胁迫响应。功能验证表明,DlDDF1是开花的负调控因子,其过表达抑制赤霉素积累并延迟开花,而KClO₃通过抑制DlDDF1、调控GA代谢及开花关键基因表达促进成花。本研究提出了KClO₃介导龙眼开花调控的分子模型,为龙眼反季成花调控与分子育种提供了理论基础。