

NG重磅 | 番茄T2T超级泛基因组解码抗逆基因
番茄在驯化过程中经历了显著的遗传瓶颈,遗传多样性与抗逆能力随之下降。相比之下,野生番茄蕴含丰富的抗病与抗逆基因资源,却长期未得到系统开发与利用。随着高质量参考基因组和属级超级泛基因组的构建,全面解析结构变异格局并精准定位关键抗性基因成为可能,为深入挖掘野生种质潜力奠定了坚实基础。
山东省精准分子作物设计育种重点实验室研究团队于2026年2月18日在国际著名期刊Nature Genetics上发表标题为“A tomato telomere-to-telomere super-pangenome empowers stress resilience breeding”的研究性论文,构建了番茄T2T超级泛基因组,系统解析其结构变异与着丝粒进化并挖掘和验证关键抗逆基因,为利用野生资源改良栽培番茄提供了重要基因组基础。

一、294份野生与栽培番茄的群体基因组学研究
本研究对236份材料进行重测序,并整合58份已发表数据,在SL6.0参考基因组基础上共鉴定约64,541,642个SNP和9,429,782个InDel,其中超过一半位于基因附近或内部。研究基于SNP将群体划分为五个类群:类群1与栽培番茄遗传距离最远;类群2-5隶属于Lycopersicon组;PIM被确定为栽培番茄最近的野生近缘种。栽培番茄核苷酸多样性较低,呈现遗传瓶颈特征,而类群3内部遗传分化显著。Fst分析表明各类群间遗传分化程度较高,整体基因交流有限,但部分物种间存在基因渗入。
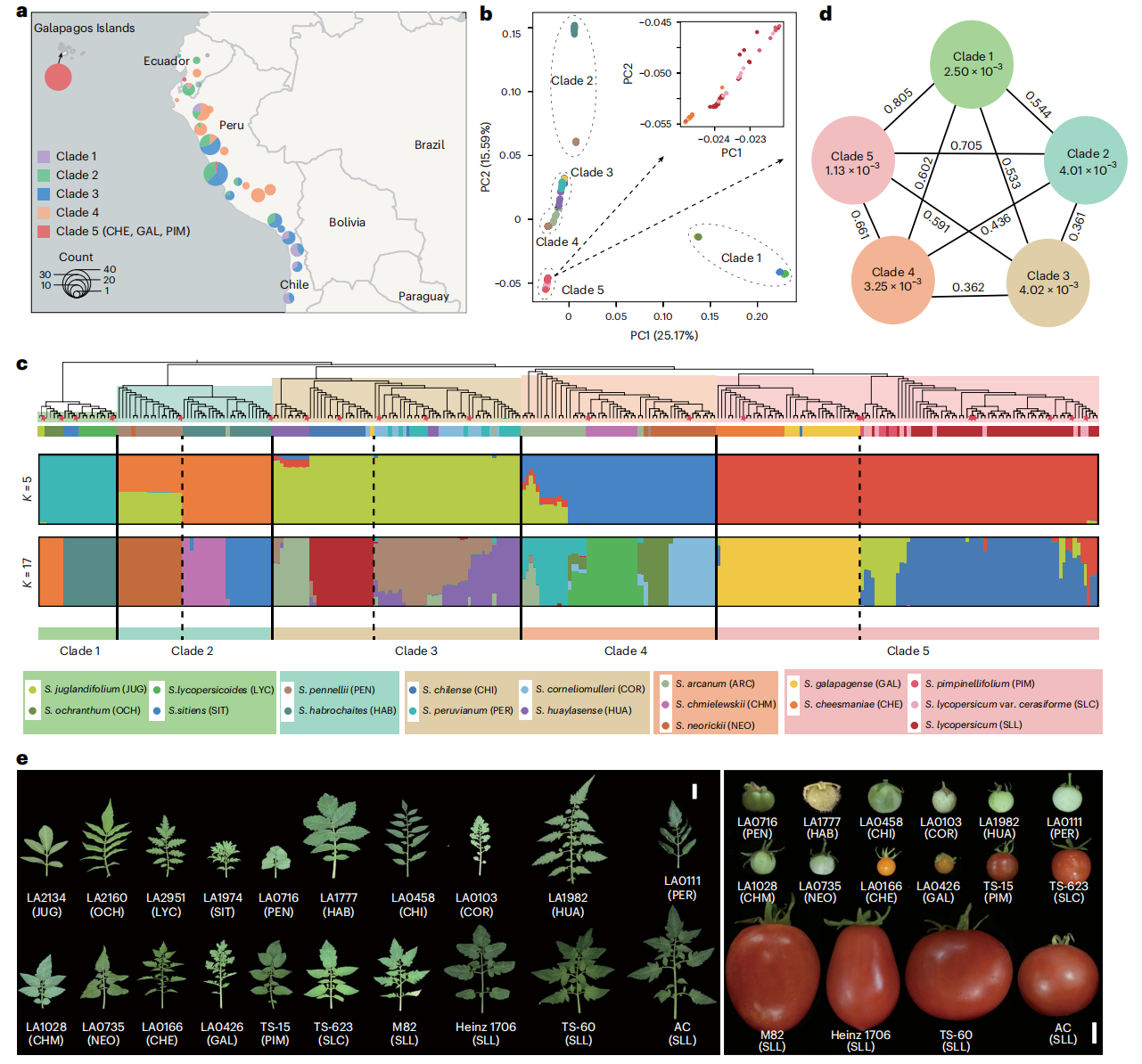
图1:野生和栽培番茄的群体分析
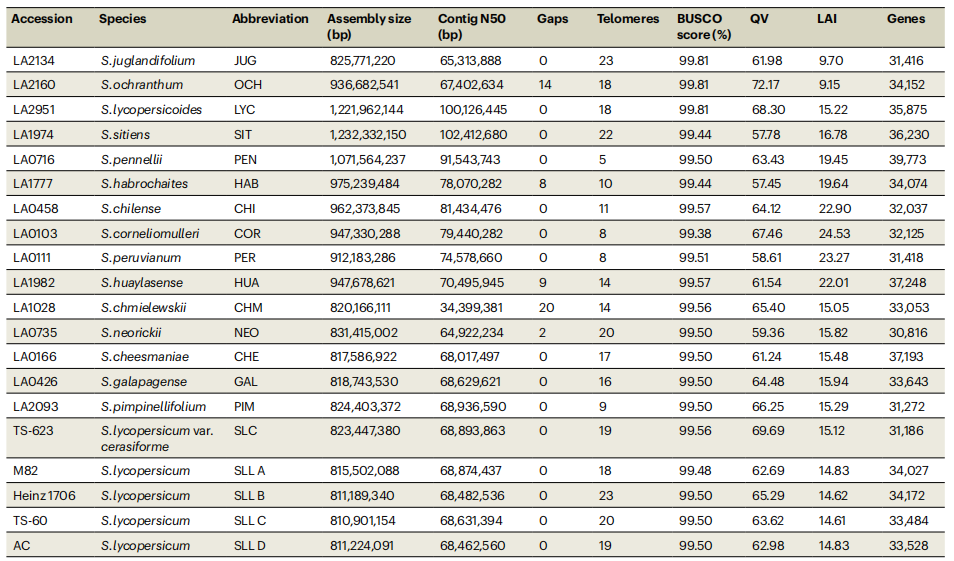
表1:20个番茄基因组组装和注释
二、番茄基因组T2T组装与系统发育分析
研究选取20个代表性番茄材料(15个野生物种及1个栽培物种的5份材料)开展T2T水平基因组组装,构建高质量番茄泛基因组参考。作者结合PacBio HiFi与ONT超长读长数据,获得15个无缺口和5个近无缺口基因组,contig N50显著优于既有版本。其中Heinz1706的SL6.0实现T2T组装,完整解析23个端粒和12个着丝粒,BUSCO达99.5%,显示出较高的完整性与碱基准确性。重复序列分析表明,转座子尤其是LTR/Gypsy元件是驱动基因组大小差异的主要因素。系统发育分析将番茄划分为五个进化类群,支持16个番茄物种的单系起源。
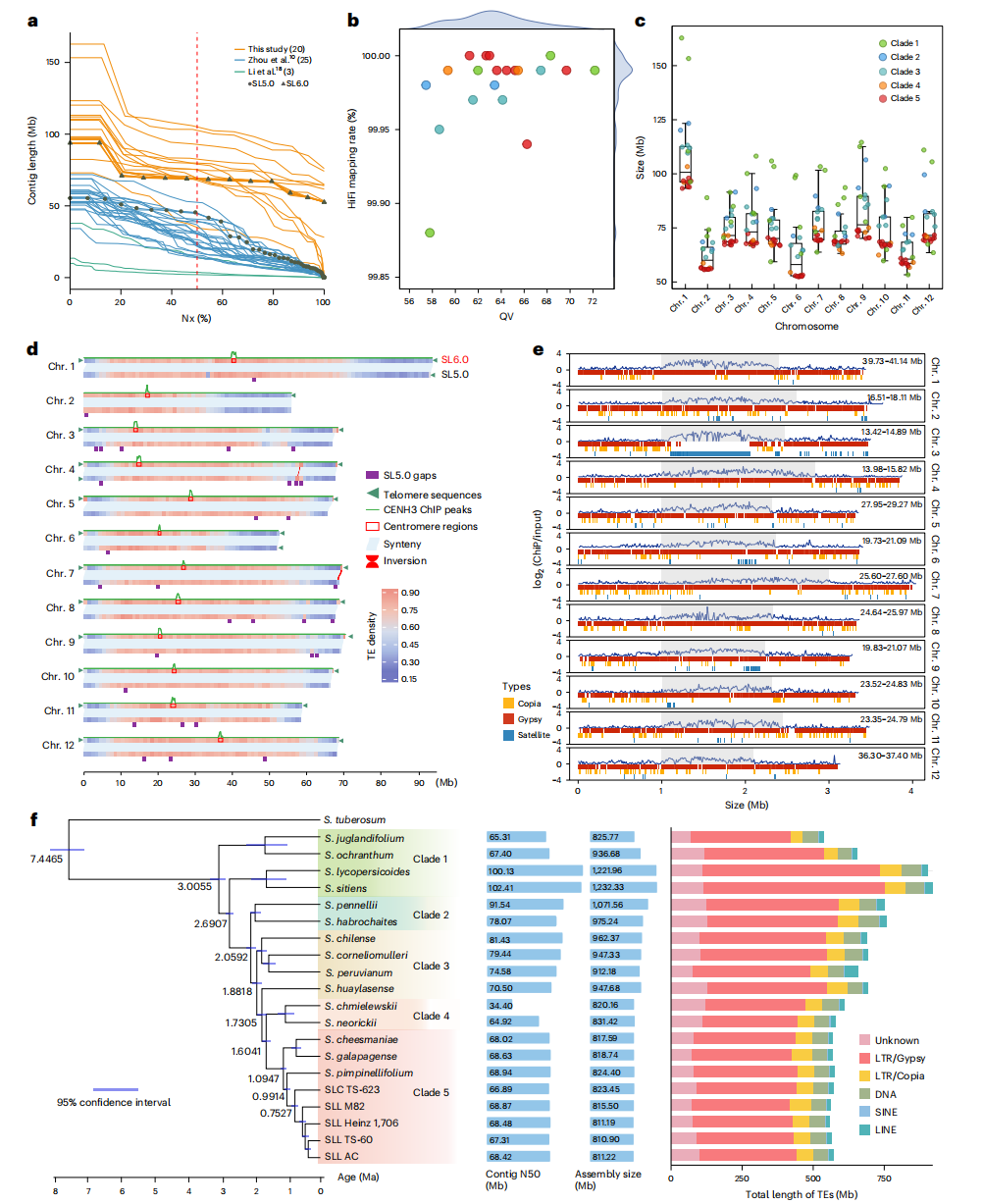
图2:野生和栽培番茄的T2T基因组组装
三、番茄泛着丝粒的多样性与进化
本研究通过对19份番茄材料开展CENH3 ChIP-seq分析并结合测序数据验证,共组装获得230个完整着丝粒。结果显示,大多数着丝粒以Gypsy类反转座子为主,部分染色体富集Sat24或Sat53卫星重复,且不同材料间着丝粒大小差异显著,野生种变异范围更广。作者推测,Sat53可能起源于45S rDNA间隔区,其形成经历由Gypsy型向Sat53富集型着丝粒转变的过程。研究还发现番茄中普遍存在着丝粒重定位现象,但多数为局部转移。
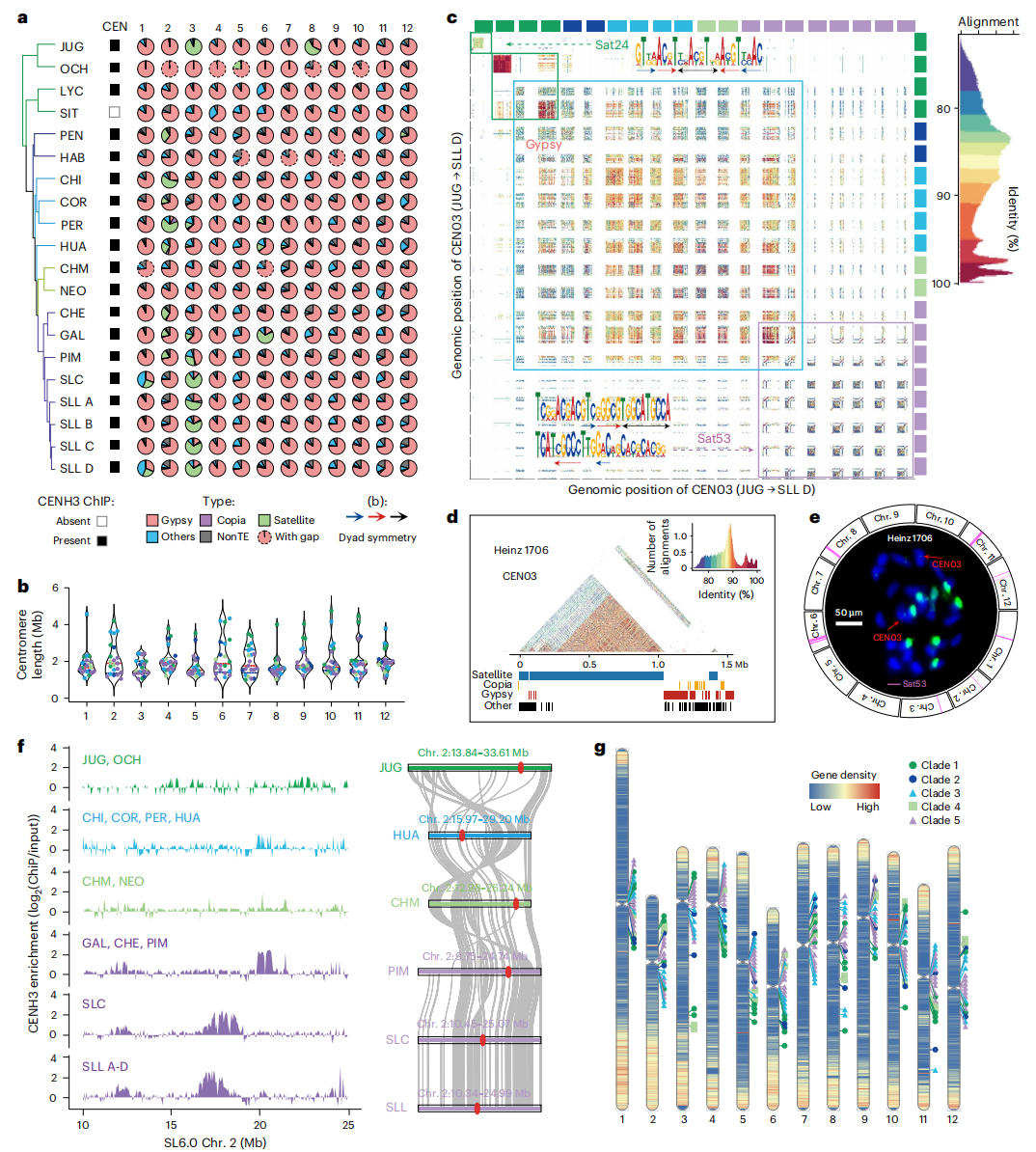
图3:番茄泛着丝粒景观及多样性
四、番茄免疫受体基因的泛基因组进化分析
基于20个高质量基因组构建番茄超级泛基因组,研究共鉴定到36,629个直系同源基因家族,其中40%为核心基因,48.2%为可变基因,2%为特有基因。可变和特有基因具有更高Ka/Ks比值和更低表达水平,说明其进化更快;功能上显著富集于防御反应、胁迫信号通路及开花调控,暗示其在环境适应中发挥重要作用。研究进一步系统鉴定NLR和RLK免疫受体基因,解析Prf/Pto抗病基因簇的茄科特异性起源及其约3300万年的进化历史,并发现两个在野生番茄中显著扩张、与灰霉病抗性相关的动态NLR基因簇,而这些基因簇在栽培番茄中发生退化或丢失,表明驯化过程可能削弱部分抗病基因资源。
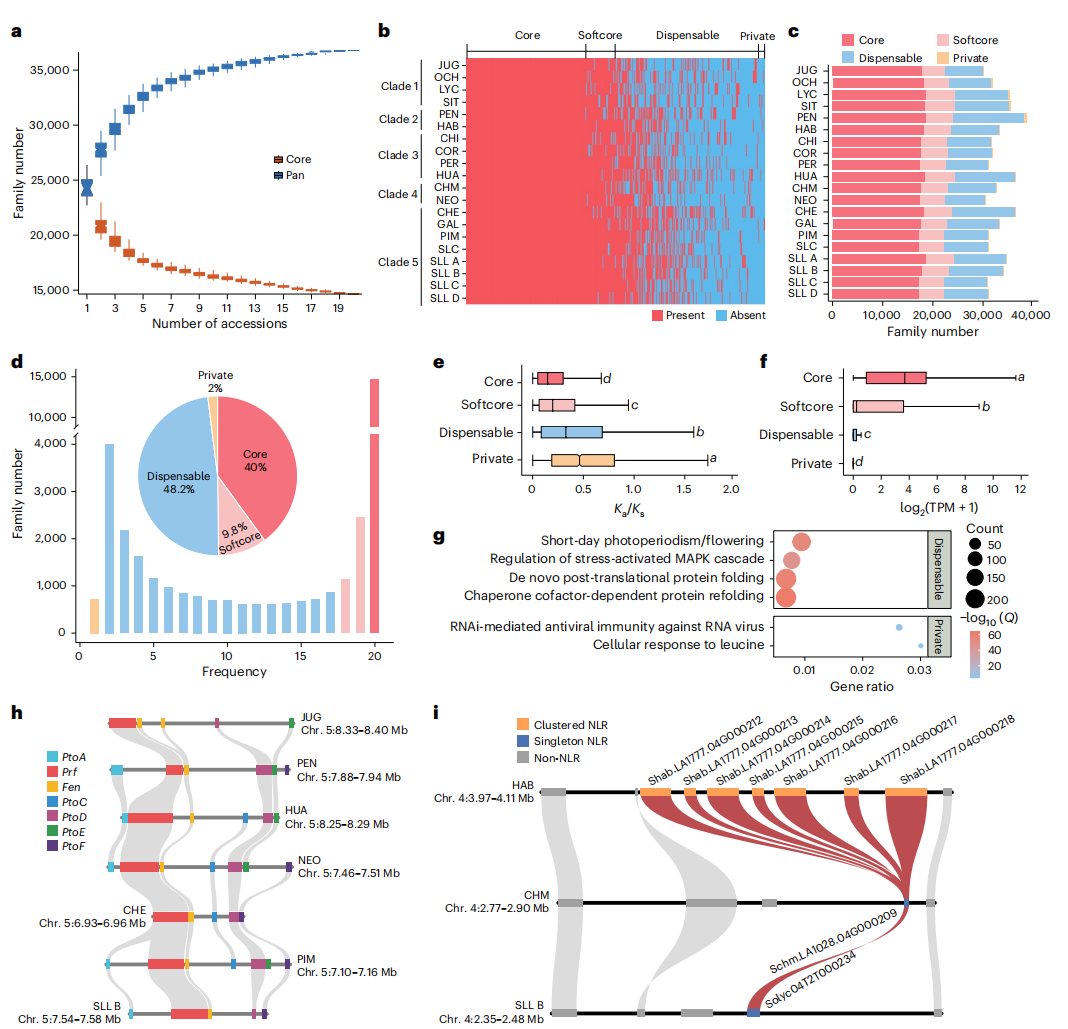
图4:20个番茄样本超级泛基因组分析
五、番茄结构变异的全面图谱
研究基于20个T2T基因组构建番茄结构变异图谱,结果显示类群1-4野生番茄存在大量基因组重排和高度分化区域,而栽培番茄保持较强共线性,反映驯化导致遗传多样性下降。未比对区域(NOTAL,nonaligned region)数量随系统发育距离递减,类群5的变异数量和长度均较低,进一步表明驯化过程中的遗传多样性丧失。作者共鉴定152,697个简单SV,包括缺失、插入、易位和倒位,其中易位和倒位平均长度更长,且在野生番茄着丝粒附近广泛存在,可能导致重组受限。整合27个已发表基因组后,共鉴定357,675个SV。未发现存在于全部46个基因组中的共享SV,且大多数SV为野生种特有。
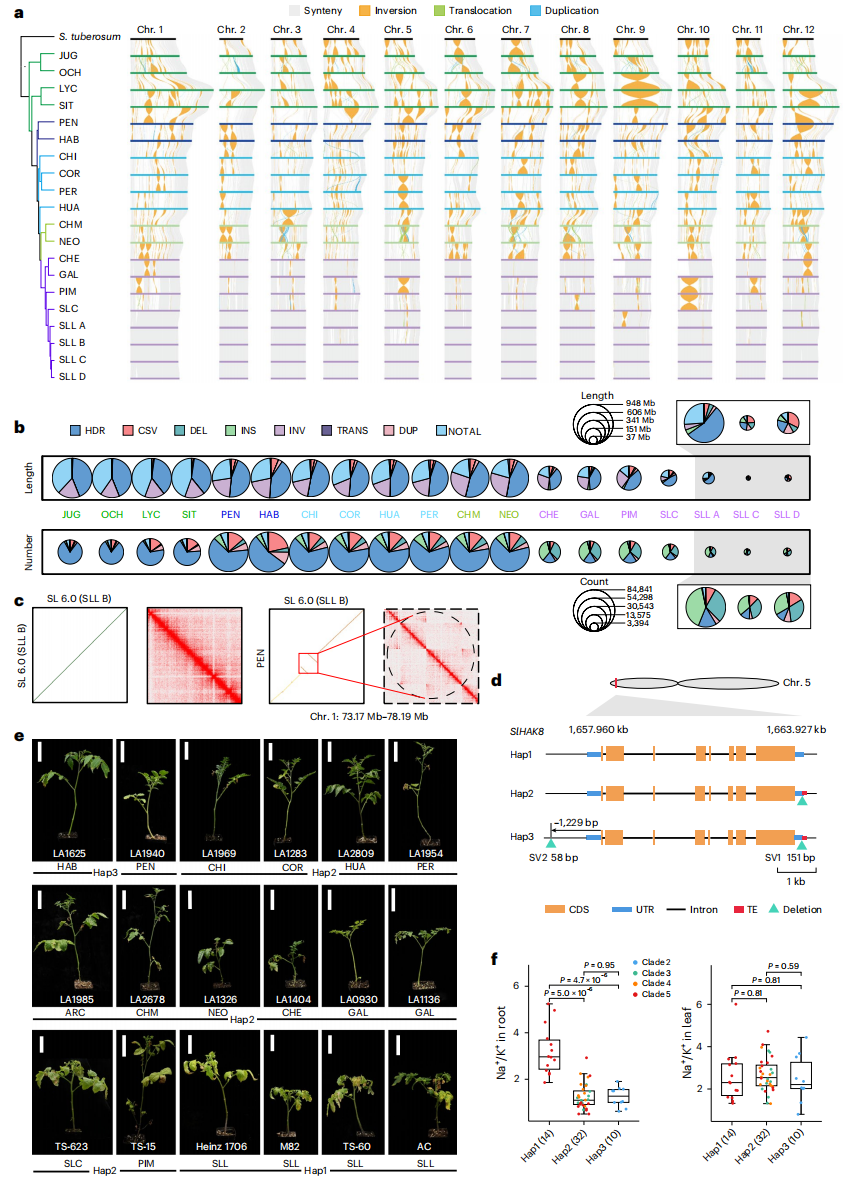
图5:番茄基因组结构变异目录揭示影响盐胁迫耐受性的功能性结构变异
六、图形泛基因组构建与SV分型分析
研究进一步构建T2T图形泛基因组,将非冗余SV整合至SL6.0,实现对294份番茄材料中101,290个SV的高效分型,并通过PCR验证其可靠性。作者筛选获得60,926个高质量SV,其中大量与基因及转座子重叠,表明转座子在SV形成中具有重要作用。功能分析表明,钾离子转运蛋白基因SlHAK8在3′-UTR和启动子区域存在两个关键SV,其不同组合形成三种单倍型(Hap1、Hap2、Hap3)。Hap2和Hap3主要存在于野生番茄,在盐胁迫下表现为根部Na?含量降低、K?含量升高、Na?/K?比值显著下降,耐盐性更强;而栽培番茄以Hap1为主,耐盐性较弱,表明在番茄驯化过程中可能丢失有利耐盐等位型。此外,本研究还鉴定出影响WRKY34和CSR等基因的SV,与番茄表型变异相关。这些关键SV及其单倍型为抗逆机制解析和分子育种提供了重要资源。
七、SV-GWAS解析灰霉病抗性基因SlGMAK
研究进一步聚焦番茄灰霉病抗性(GMR)的遗传基础,对209份番茄材料进行系统分析,发现不同类群间抗性差异显著,其中OCH、GAL、CHE及部分HAB材料表现出较强抗性。连锁不平衡分析显示,仅约4%的SV与SNP或InDel存在较强连锁,表明大量SV相关遗传变异尚未被解析。作者基于SV基因型开展GWAS,共鉴定18个显著SV-QTL,其中9号染色体上937 bp缺失位点关联信号最强。该缺失位点与抗性表型高度相关,位于LRR-RLK基因SlGMAK下游。功能验证表明, SlGMAK正向调控灰霉病抗性,937 bp缺失可抑制其表达并与抗性性状共分离。
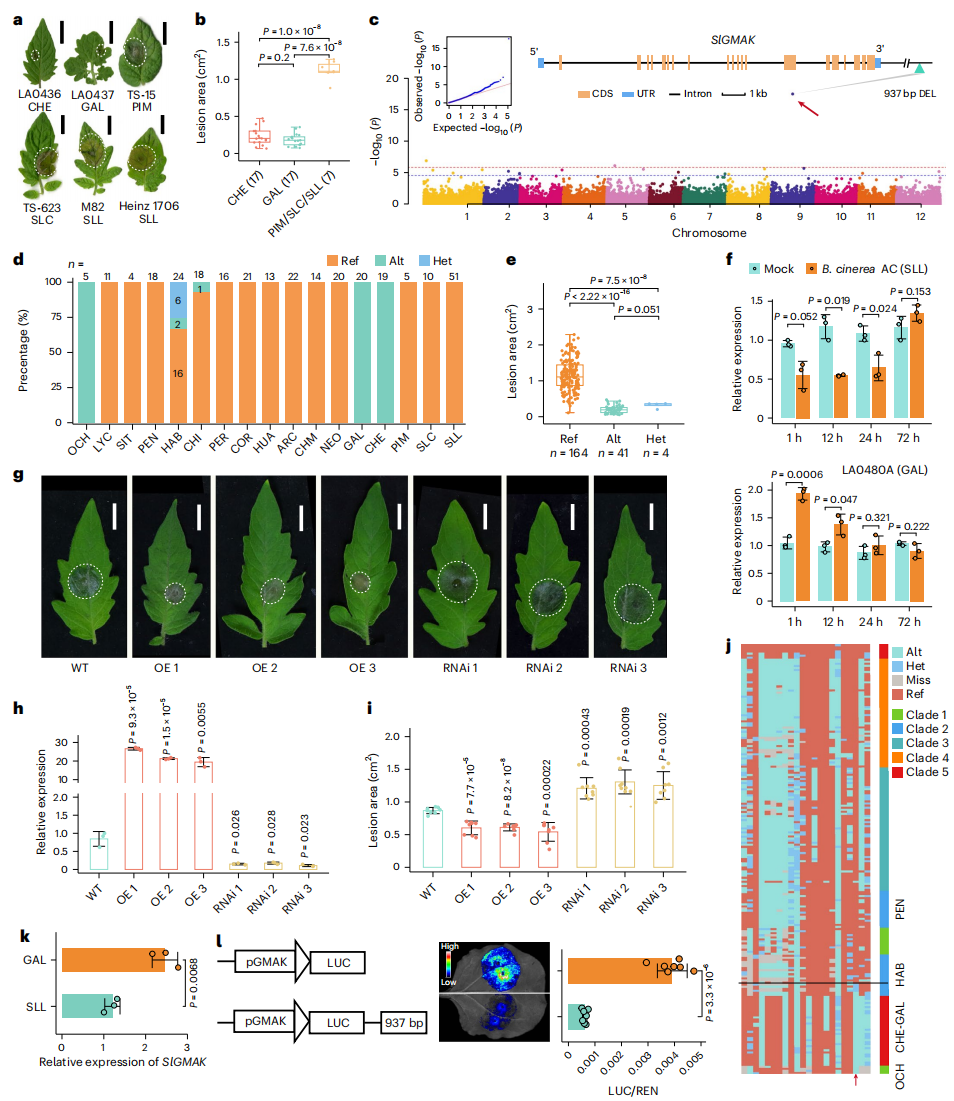
图6:SV-GWAS鉴定出一个GMR基因
结语
本研究构建了高质量番茄T2T超级泛基因组,整合野生与栽培资源,系统解析了全基因组结构变异及着丝粒特征,揭示了番茄类群的遗传结构与进化关系。基于图形泛基因组开展SV-GWAS,克隆并验证了灰霉病抗性基因SlGMAK,并挖掘出多个与抗性和耐盐性相关的关键变异位点。同时构建泛NLRome和泛RLKome,拓展了抗病基因资源。研究为恢复驯化过程中丢失的遗传多样性、推动番茄抗逆分子育种提供了重要基因组基础与理论支撑。