

NC重磅 | 茄子泛基因组揭示果实颜色遗传基础
茄子(Solanum melongena L.)是全球重要的茄科蔬菜作物,产量仅次于马铃薯和番茄,但其起源与驯化历史仍存在争议。近年来基因组资源的积累推动了对茄子遗传多样性和重要农艺性状遗传基础的研究。然而,现有研究多依赖单一参考基因组或SNP变异分析,难以全面捕获种内丰富的结构变异与遗传多样性。
广东省农业科学院蔬菜研究所研究团队于2026年2月23日在国际著名期刊Nature Communications上发表了标题为“Sub-pangenome analysis reveals structural variants associated with fruit color and bacterial wilt resistance in eggplant”的研究性论文,通过构建茄子泛基因组并结合群体基因组分析,鉴定到与果实颜色和青枯病抗性相关的重要结构变异,为解析茄子性状形成机制及其遗传改良提供了重要的基因组资源。
一、基因组测序、群体结构与遗传多样性
本研究对226份茄子种质资源进行了重测序,包括219份栽培茄子、2份野生近缘种、4份红茄和1份野生茄子,主要分布在东亚和东南亚地区。研究共获得5.50 TB测序数据,平均测序深度为20.87×,并以华南育种系“S076”为参考基因组进行比对,鉴定出3,698,811个SNP和349,227个InDel。基于SNP进行PCA和群体结构分析,将材料划分为四个群体(C1-C4):C1主要来自东南亚,C2来自南美和欧洲,C3主要分布于中国各省,C4主要来自中国华南地区。结果表明茄子群体结构与地理分布密切相关。核苷酸多样性分析显示C1和C2群体的遗传多样性高于中国的C3和C4群体,且C2与C4之间遗传分化最大,而C3与C4遗传距离最近。
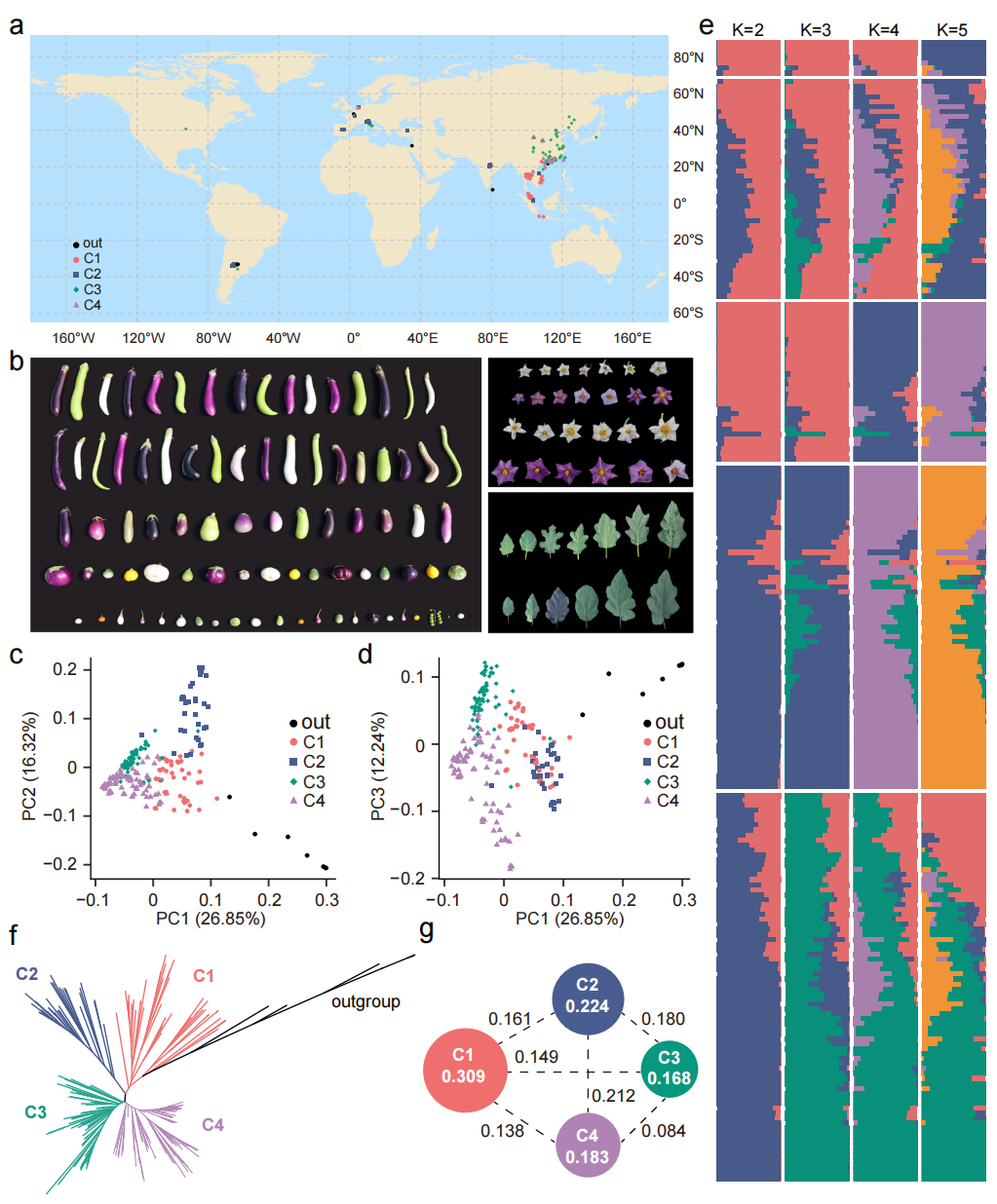
图1:226份茄子材料的地理分布及基因组多样性
二、茄子基因空间的亚泛基因组
本研究结合226份茄子材料的重测序数据及4个已发表基因组,筛选出17个代表不同系统发育分支的材料,构建茄子基因空间的亚泛基因组。其中对11份材料进行从头测序,获得PacBio HiFi(30.18×)和Hi-C(103.79×)数据,组装得到染色体水平基因组,contig N50为38.73-59.67 Mb,组装大小1.14-1.20 Gb,BUSCO完整度均超过98.70%,表明组装质量较高。基因组注释显示每个基因组含68.79%-79.85%的重复序列和33,620-36,174个蛋白编码基因。基于17个基因组开展泛基因组分析,随着基因组数量增加,泛基因家族数量逐渐增加并在17个基因组时趋于稳定。最终鉴定出15,406个核心基因家族、3,657个软核心基因家族、17,889个可变基因家族和483个特有基因家族。核心基因表达水平较高且Ka/Ks值较低,而可变基因家族富集于防御反应相关GO功能,可能与茄子抗病性及表型多样性形成有关。
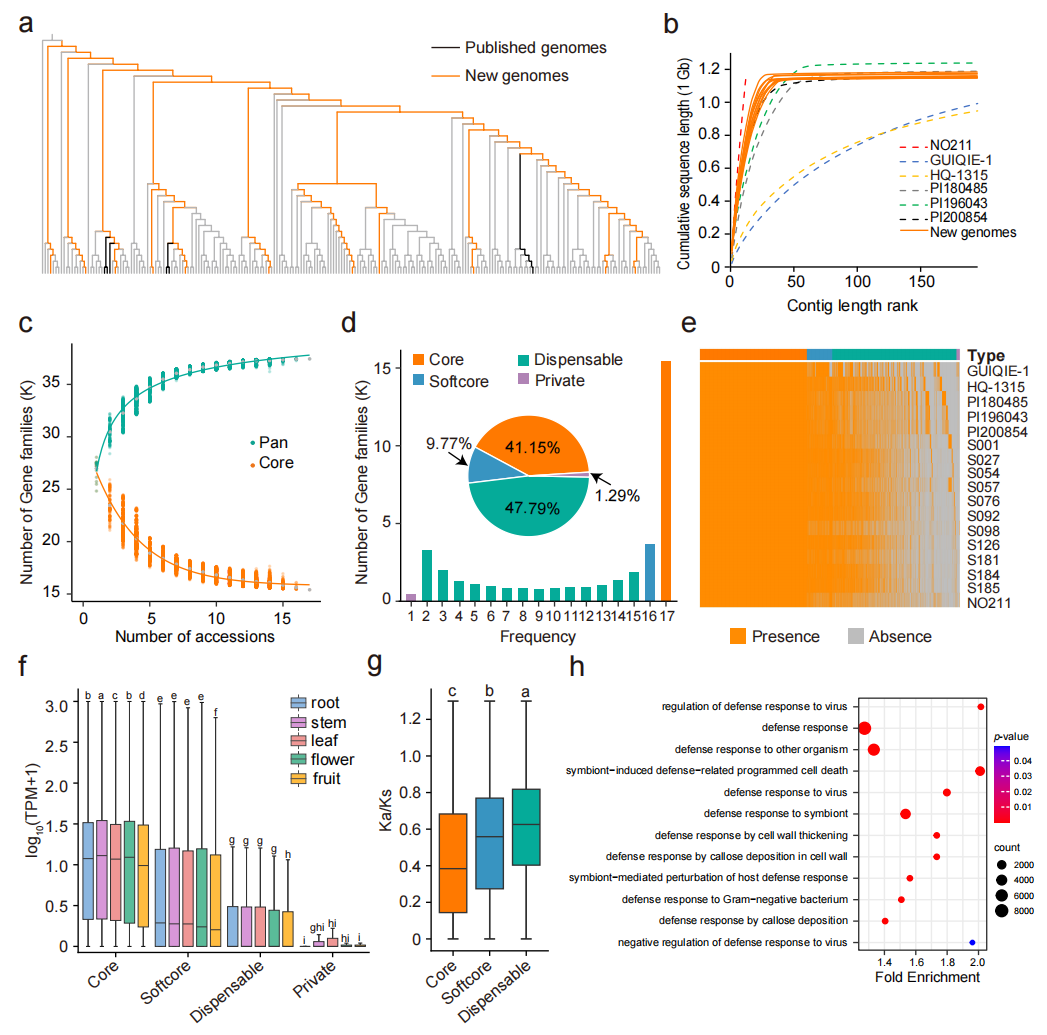
图2:17份茄子材料的泛基因组分析
三、茄子结构变异的群体特征、适应度及功能效应
本研究基于16个茄子基因组(其中10个为本研究测序)系统鉴定长度大于50 bp的结构变异(SV),并以S. insanum和S. violaceum作为外群进行比较分析。通过与参考基因组“S076”比对,共检测到156,540个SV,包括插入、缺失、易位和倒位四种类型;整合分析获得76,481个唯一定位SV,其中约60%为材料特异变异。群体频率分析表明,大多数SV以低频形式存在,其派生等位基因频率显著低于SNP,说明SV整体上对个体适应度具有更强的负效应。功能分析显示,SV在基因区域显著缺乏,仅少量直接影响编码序列。表达分析进一步发现,含SV的基因表达变异显著高于不含SV的基因,并鉴定出93个SV显著影响表达的基因。
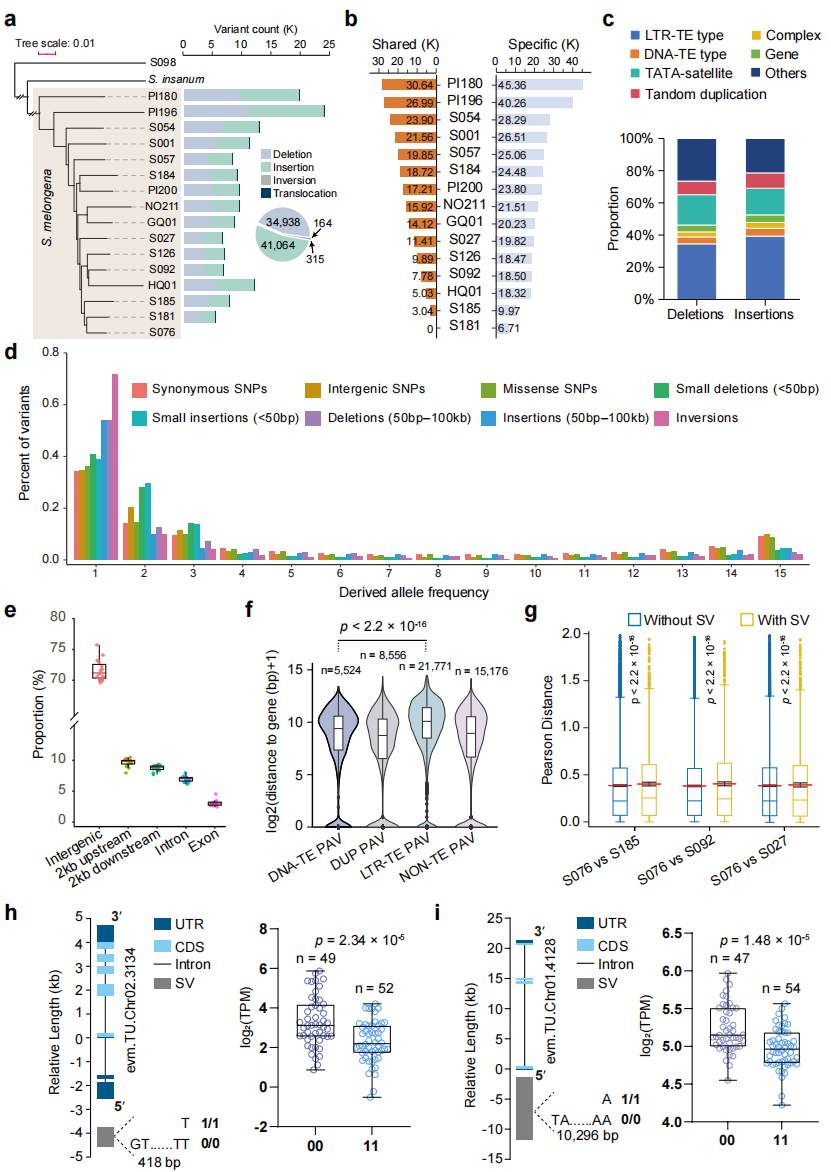
图3:茄子中的结构变异
四、基于图形结构的茄子亚泛基因组
本研究基于226份茄子材料的重测序数据鉴定大于50bp的缺失和插入并与组装基因组检测到的SV整合,共获得187,412个非冗余结构变异。以参考基因组S076为骨架,构建图形泛基因组并对所有材料进行SV基因型鉴定,结果显示大多数SV在群体中以低频存在。由于样本主要来自东亚、东南亚和欧洲,该资源被定义为茄子“亚泛基因组”。此外,基于16个栽培茄子基因组利用PGGB构建无参考泛基因组图,以减少参考偏倚。该图包含3180万个节点和4390万条边,总序列长度1.96Gb,并解析出31,793个SV、2,045,873个InDel和6,939,066个SNP,为解析结构变异对茄子表型变异的功能影响提供了重要资源。
五、与茄子果色显著关联的10号染色体倒位
本研究在201份栽培茄子材料中开展GWAS,将果色划分为紫色和非紫色两类,在10号染色体上检测到一个跨度超过10Mb的显著关联区域,其中包含多个花青素合成相关基因(如SmANS、Sm3GT、SmMYB1和SmPAL)。进一步分析发现,该区域与一个约12.4Mb的大型倒位结构变异一致,且显著位点富集于倒位断点附近。群体基因型分析表明,携带该倒位的材料中96.52%为紫色果实,说明该倒位与紫色表型高度相关,但并非唯一决定因素。表达分析显示,在倒位区域相关基因中仅SmMYB1表现出显著差异表达,并在绿色或白色果实材料中存在6bp缺失。结果表明SmMYB1可能是影响茄子果色的重要候选基因,并可能与该倒位共同参与紫色果实形成的调控。
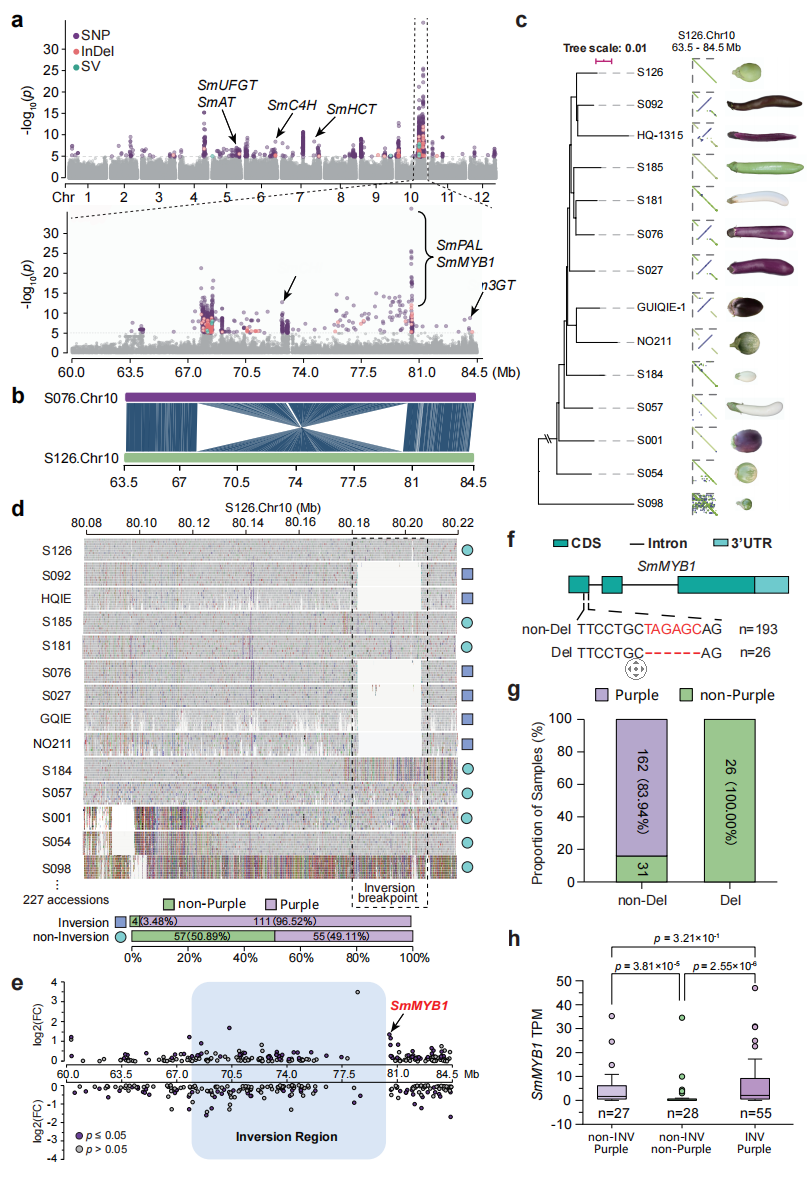
图4:一个高频的大型倒位(约12.4 Mb)与茄子果实颜色显著相关
六、12.4 Mb倒位与遗传多样性及单倍型结构
进一步分析了该倒位的群体遗传特征,结果表明该倒位在群体中的等位基因频率约为50.44%,主要分布于中国来源的C3和C4群体,提示其可能在中国茄子驯化或育种过程中频率上升。群体遗传分析表明,不携带倒位的材料在Chr10上的核苷酸多样性(π)约为倒位群体的两倍,且倒位区域Fst和Tajima’s D显著升高,说明该区域存在明显遗传分化并可能受到选择。系统发育分析显示,携带倒位的材料形成独立分支并共享单一单倍型。进一步分析发现,该倒位区域形成约6Mb的连锁不平衡(LD)区块,说明该区域重组受到明显抑制。结合其与果色的强关联关系,推测该倒位可能起源于denovo突变,并在茄子栽培与育种过程中受到人工选择而在群体中维持较高频率。
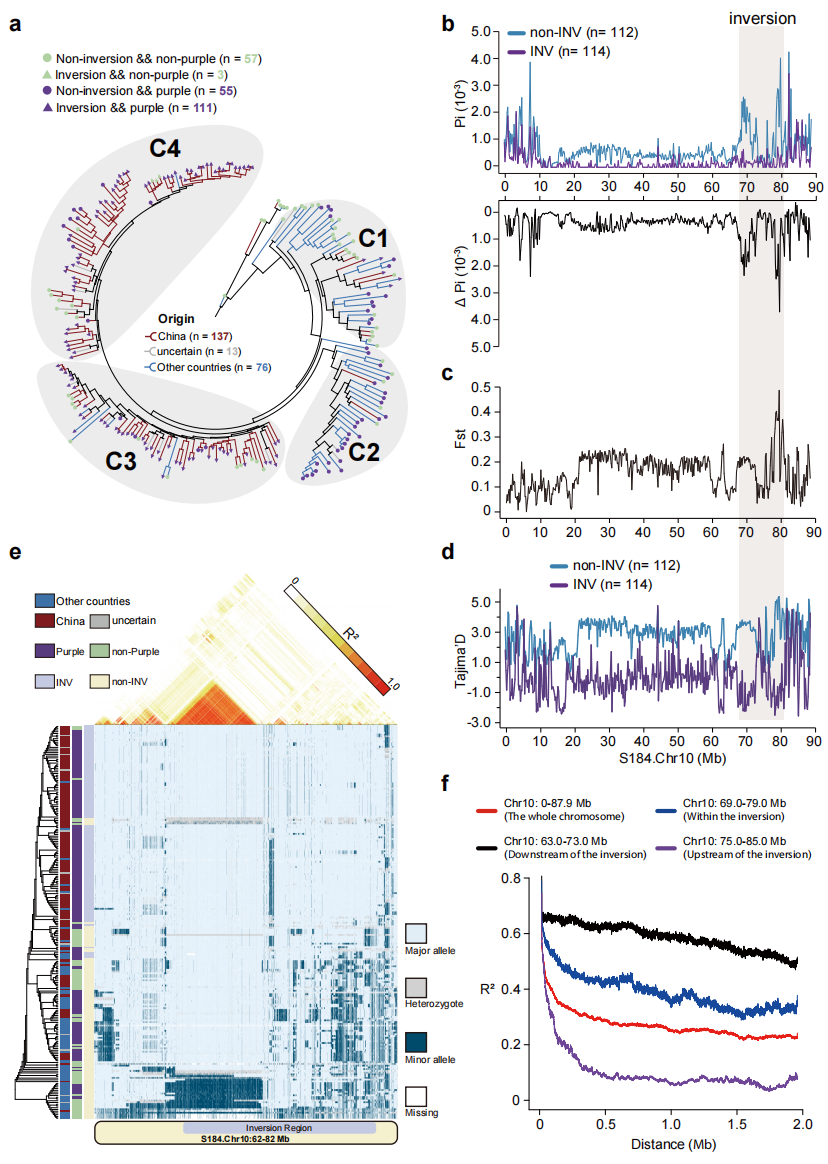
图5:12.4 Mb倒位区域的遗传多样性降低及扩展的单倍型结构
七、与青枯病抗性相关的基因组位点
为解析茄子青枯病(BW)抗性的遗传基础,本研究对197份材料接种青枯雷尔氏菌Ralstonia solanacearum并开展GWAS分析,在 Chr02、Chr03、Chr04 和 Chr05 上检测到显著关联信号。其中,Chr04上的关键SNP位于 SmCYP82D47 基因第一外显子,可产生提前终止密码子并导致蛋白缺失大部分细胞色素P450保守结构域,从而显著影响抗病性。Chr05区域的关联信号主要集中在 EPS1 同源基因及邻近的 Roq1-like 抗病基因附近。比较基因组分析表明该区域存在明显的基因拷贝数变异,并鉴定出两个结构变异标记 SVe(EPS1扩增)和 SVr(Roq1-like扩增)。联合Chr04位点分析表明,这些标记与青枯病抗性显著相关。进一步通过VIGS实验验证 SmCYP82D47、SmEPS1 和 SmRoq1 在抗病过程中发挥重要作用,为茄子抗青枯病分子育种提供了重要候选基因和分子标记。
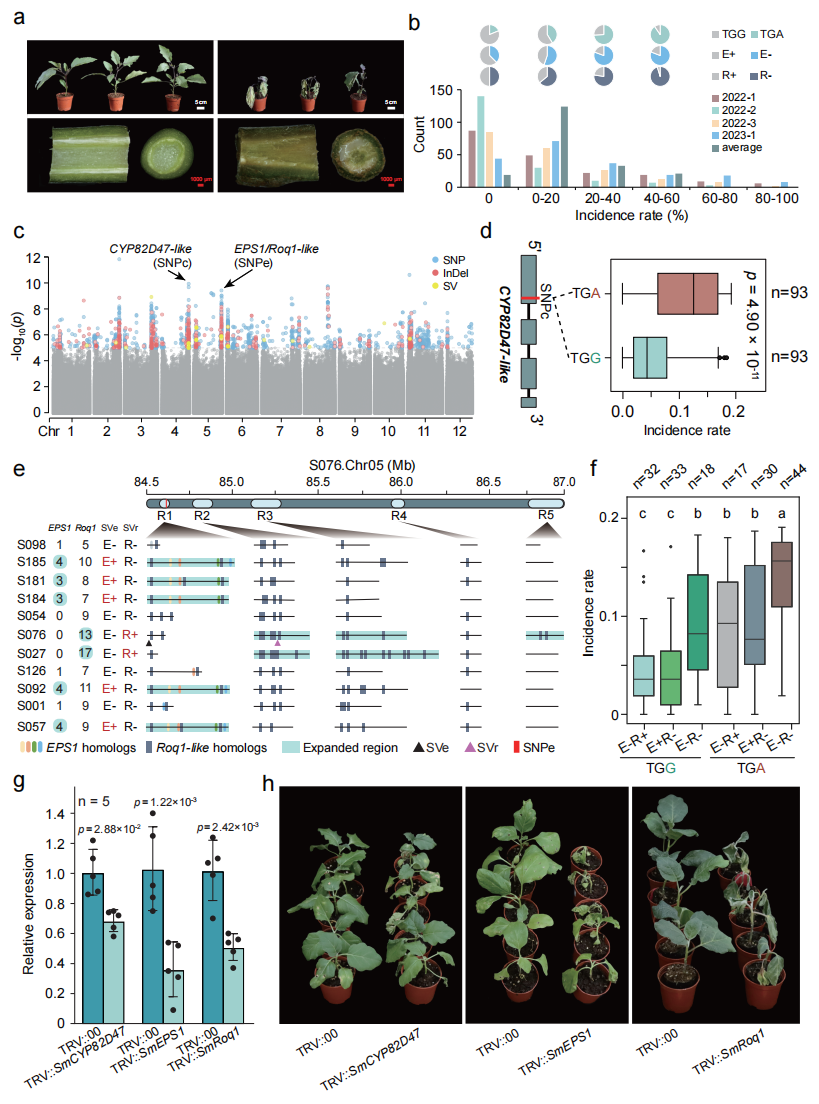
图6:通过GWAS鉴定的青枯病抗性候选位点
结语
本研究以226份茄子材料为基础,结合多基因组组装与重测序数据构建了茄子亚泛基因组,并系统解析了群体结构、遗传多样性及结构变异特征。结果表明,茄子群体遗传分化与地理来源密切相关,结构变异在重要农艺性状形成中发挥关键作用。其中,10号染色体上约12.4 Mb的大型倒位与果实紫色性状显著相关;同时鉴定到多个与青枯病抗性相关的候选基因及分子标记。该研究为阐明茄子重要性状的遗传基础及开展分子育种提供了重要的基因组资源。