

Nature重磅 | 裂腹鱼染色体融合与重新二倍化
全基因组复制(WGD)在真核生物演化中具有重要意义,通过产生大量重复基因,为谱系分化及新功能演化提供了遗传基础。尽管WGD通常发生于脊椎动物早期演化并为其复杂性发展奠定了遗传基础,但其后续重新二倍化过程至今仍不完全明确。裂腹鱼(Schizothoracinae)由于其多倍体特性和演化历史,成为了研究这一过程的理想模型。
西南大学徐洛浩教授、刘海平教授与Axel Meyer教授联合团队在国际著名期刊Nature上发表标题为“Chromosomal fusions trigger rediploidization of autopolyploid genomes”的研究性论文,通过基因组分析揭示了裂腹鱼类群的复杂倍性演化过程,重点探讨了染色体融合、重新二倍化以及基因组结构重塑对物种演化的驱动作用。

一、裂腹鱼倍性确认与基因组组装
本研究聚焦裂腹鱼类群的倍性分型与基因组组装。研究基于覆盖11个裂腹鱼属的短读长测序数据,通过K-mer频率分布分析其多倍体类型。结果表明,大多数裂腹鱼为同源四倍体,同时还识别出二十倍体和六倍体物种,这说明该类群经历了复杂的倍性演化。研究进一步选取拉萨裂尻鱼(Schizothorax younghusbandi)进行高质量单倍型基因组组装。通过构建亲本-子代三联体,并结合PacBio HiFi、ONT超长读长和Hi-C数据,研究采用读段分箱与重新组装策略,最终获得了包含 90 条染色体的单倍型分型基因组。
二、不对称染色体融合
研究发现,拉萨裂尻鱼的染色体数为90条,少于同源四倍体理论预期的100条。通过与外群物种进行基因组共线性比较,研究确认染色体数目的减少主要由五对染色体融合造成,而并非染色体丢失。进一步分析表明,这些融合具有不对称性:融合仅发生在四条同源染色体中的两条上,另外两条仍保持未融合状态,并经FISH实验验证。该现象表明,多倍化后的裂腹鱼基因组中,同源染色体已开始分化为融合拷贝和未融合拷贝,并可能沿不同的进化轨迹发展。同时,不对称染色体融合也说明拉萨裂尻鱼基因组可能正处于重新二倍化过程中。
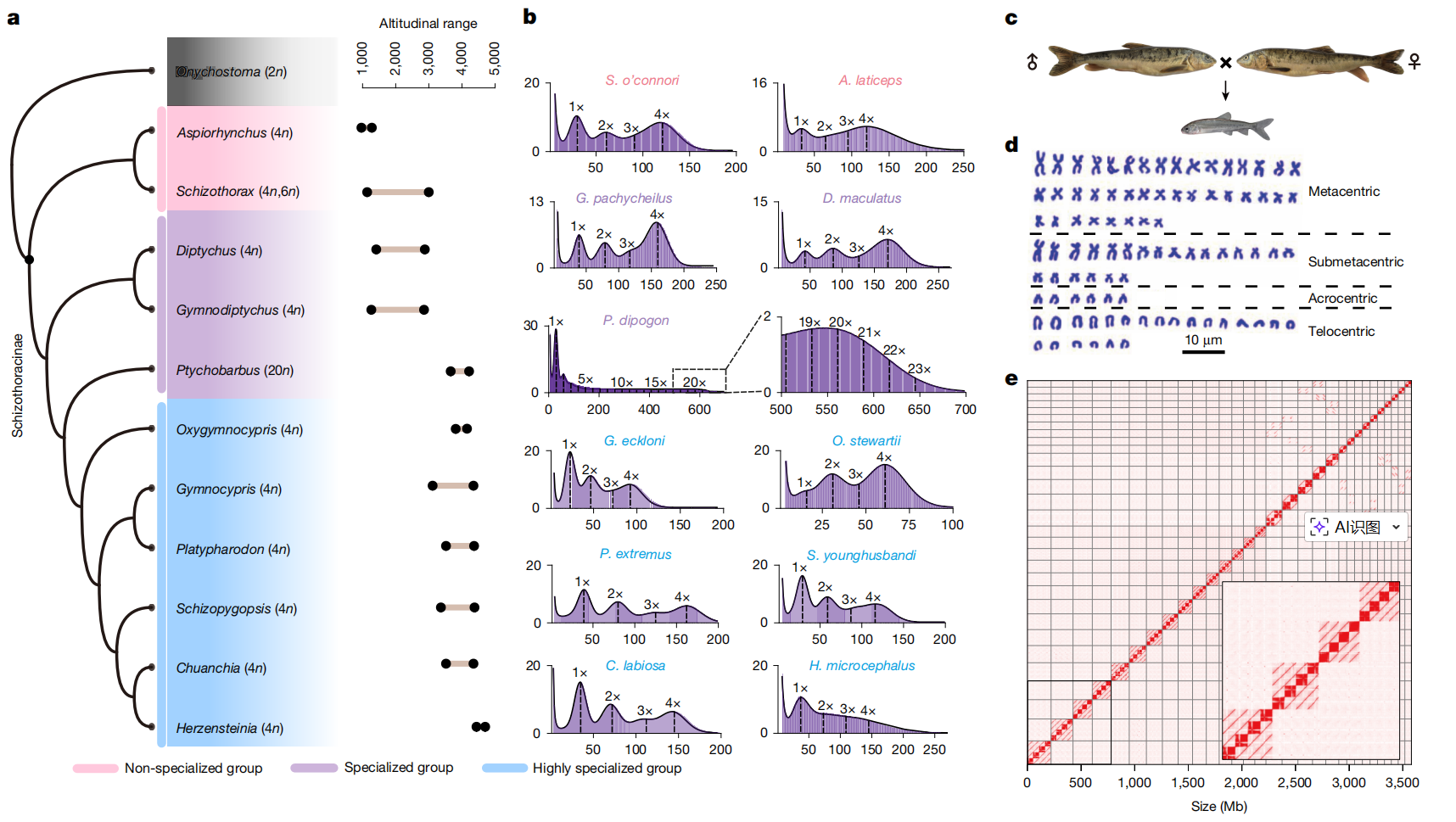
图一:裂腹鱼属均为同源多倍体
三、不同步重新二倍化的起始
研究通过比较同源染色体四联体的序列分歧(Ks值),发现未融合四联体分歧较低,主要保持四体遗传;而发生不对称染色体融合的四联体分歧较高,说明融合可能改变了减数分裂中的染色体配对方式,推动部分区域向二体遗传转变。其中,染色体19和22的分歧最高,可能代表最早一波重新二倍化;染色体11和14分歧较低,说明其仍处于早期阶段。等位基因分析也支持二体遗传与四体遗传在同一基因组中共存,表明拉萨裂尻鱼的重新二倍化是一个分阶段、不同步推进的过程。
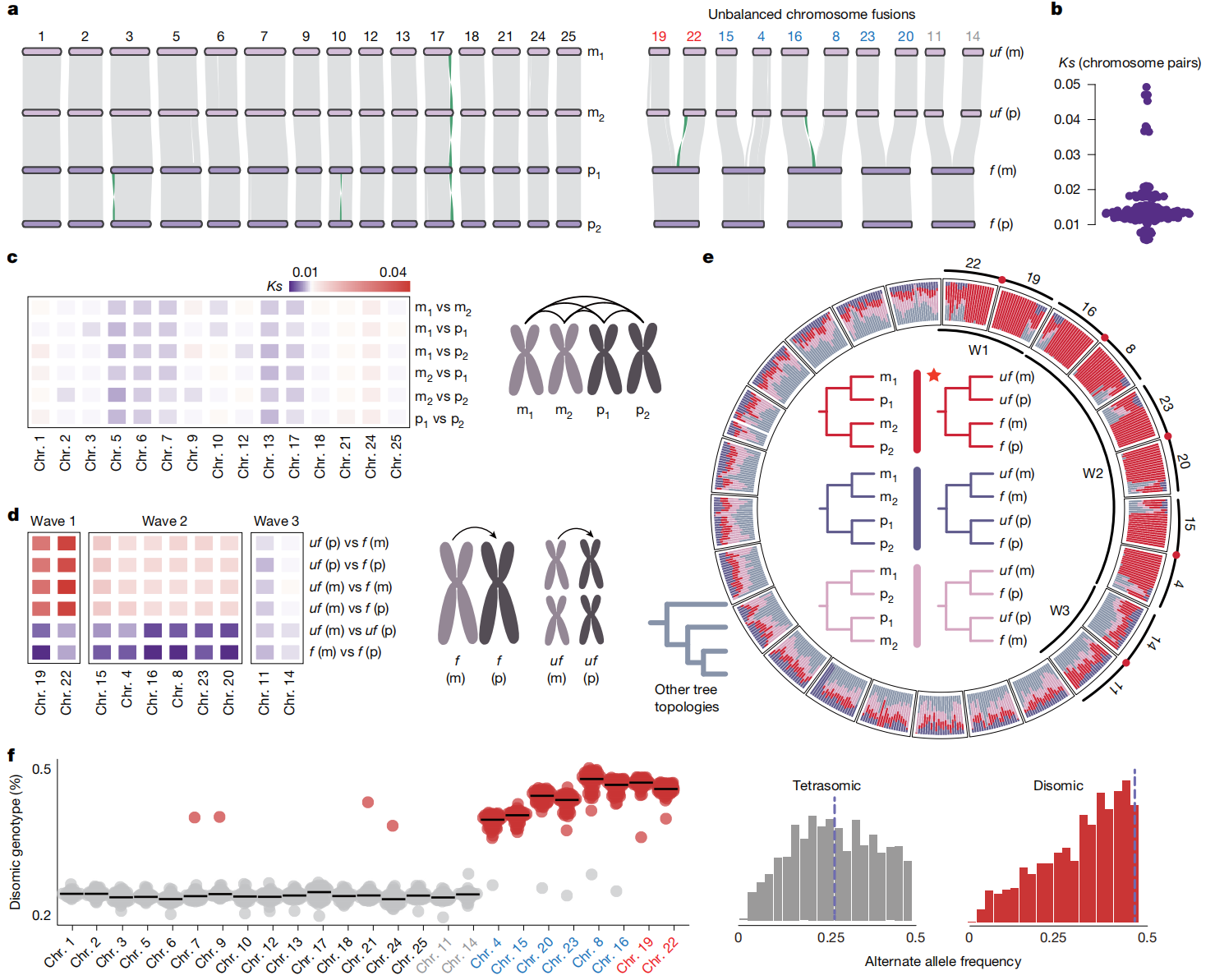
图二:拉萨裂尻鱼基因组中的三波重新二倍化
四、重新二倍化始于融合位点
研究进一步发现,二体基因型主要集中在染色体融合位点附近,而远离融合位点的染色体臂和端粒区域更多保留四体遗传,说明重新二倍化可能从融合位点开始,并逐步向外扩展。滑动窗口分析显示,融合区域附近的序列分歧更高,提示这些区域较早发生分化。分子钟估算表明,染色体19和22的融合及第一波重新二倍化约发生在3000万年前,第二波重新二倍化约始于1000万至 2000万年前,染色体融合可能是启动裂腹鱼重新二倍化的重要因素。
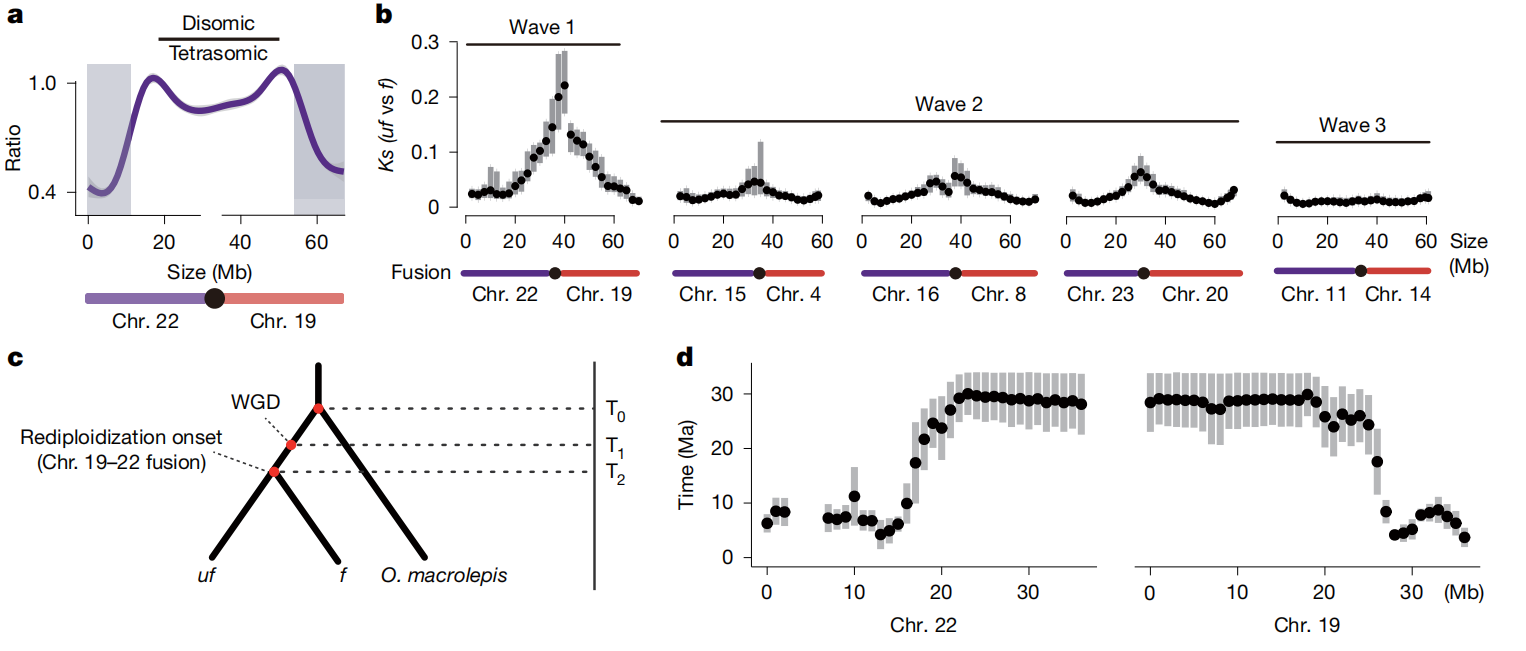
图三:重新二倍化始于染色体融合位点
五、染色体行为
研究从细胞遗传学层面观察了拉萨裂尻鱼的染色体行为,发现其减数分裂染色体中同时存在环状和链状结构,分别与中心着丝粒染色体和端着丝粒染色体配对有关。着丝粒序列分析显示,端着丝粒染色体主要使用Cen263重复序列,其他染色体则主要使用Cen254。进一步探针杂交结果支持四价配对的存在,并显示长链结构可能由融合染色体与未融合染色体共同配对形成。此外在染色体3和17中还发现着丝粒重新定位事件,说明着丝粒倒位可能参与基因组结构重塑,并与重新二倍化过程相关。
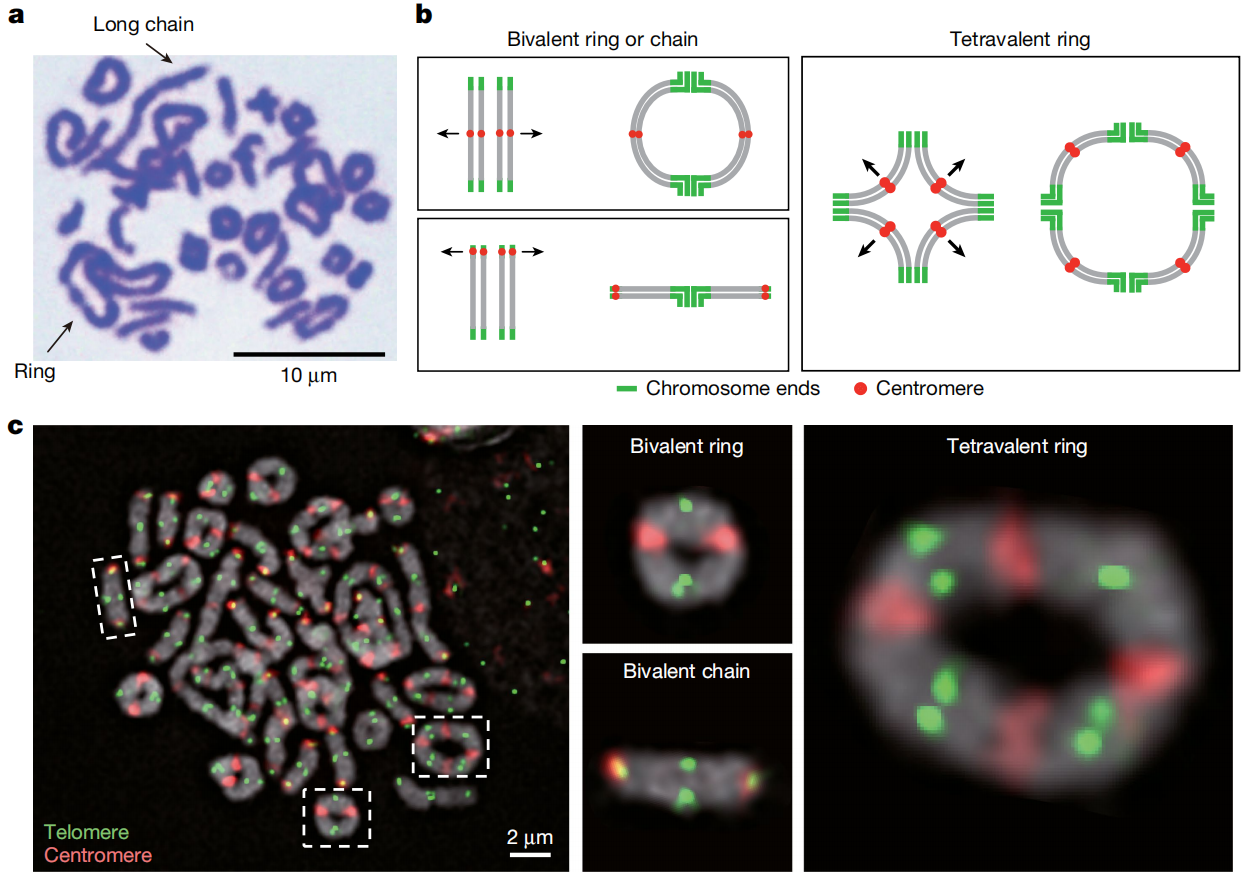
图四:减数分裂中期 I 的染色体行为
六、不对称同源基因演化
研究进一步分析了裂腹鱼同源基因的丢失与表达分化。结果显示,虽然多数基因仍保留四个拷贝,但部分基因已丢失至三个或两个拷贝,且这种丢失主要发生在融合染色体上。RNA测序结果表明,未融合拷贝(uf)的表达水平整体高于融合拷贝(f),尤其在第一波重新二倍化染色体中最为明显。部分同源基因在不同组织中表现出相反的表达偏向,说明其可能发生功能分化。低温应激实验显示,uf与f拷贝通常同步响应环境变化,说明二者仍可能保持协同调控,并可能参与高原环境适应。
七、共同的自体多倍体化
研究追溯了裂腹鱼类群多倍体化的共同起源。结果发现,不同裂腹鱼物种的染色体19和22均表现出类似二倍体的遗传模式,并在Hi-C数据中显示明显互作信号,说明这两条染色体的融合很可能发生在裂腹鱼共同祖先中。弧唇裂腹鱼(Schizothorax curvilabiatus)的单倍型基因组也呈现出相同的融合信号,进一步支持裂腹鱼类群在物种分化前经历了共同的自体多倍体化事件。其他染色体融合仅出现在部分高度特化类群或拉萨裂尻鱼中,说明重新二倍化是在不同谱系中分阶段且不同步推进的。
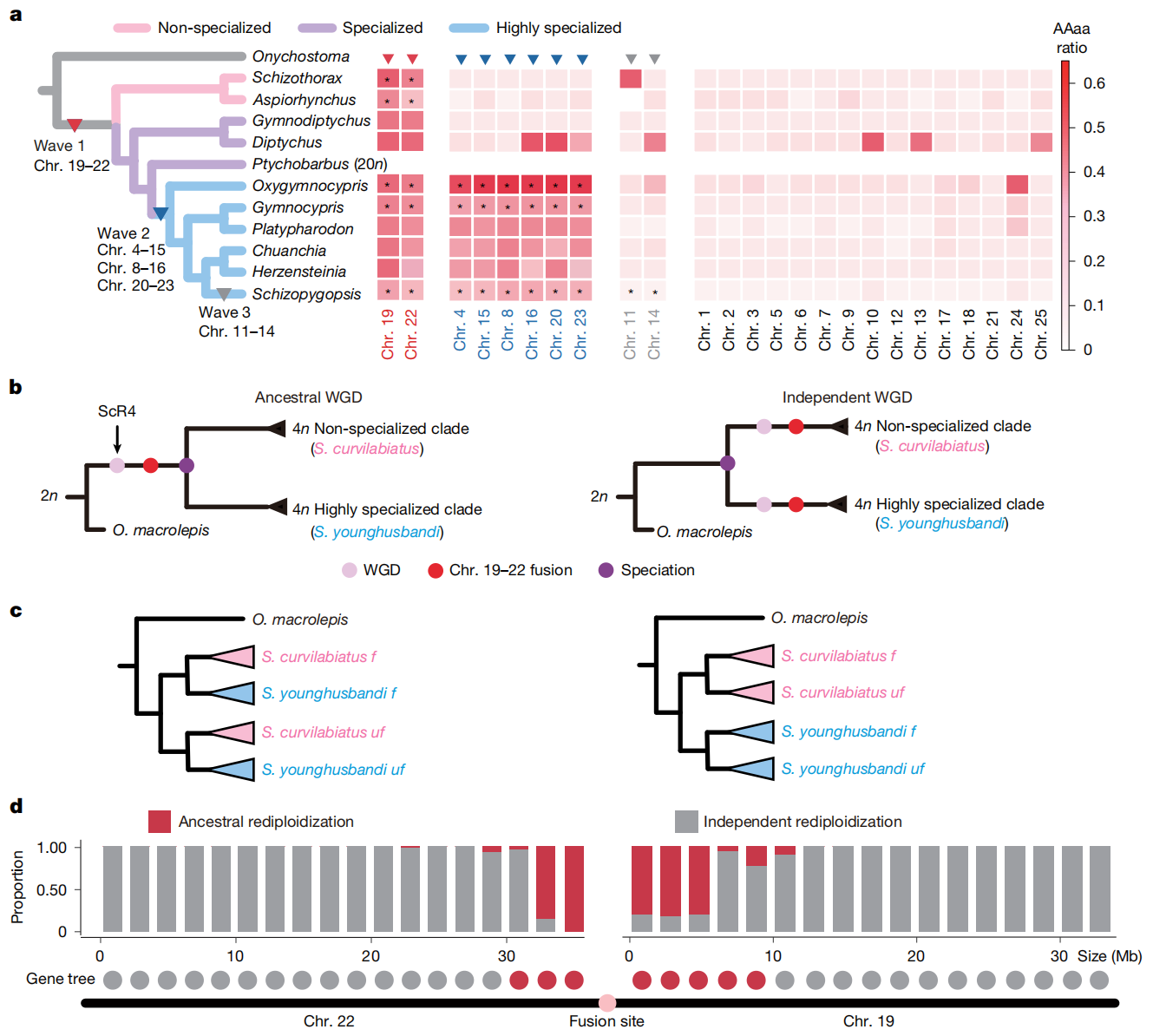
图五:裂腹鱼类群中的一次祖先全基因组复制事件
结语
本文研究了裂腹鱼类群的倍性演化及其基因组结构,确认大多数物种为同源四倍体,同时识别了二十倍体和六倍体物种,揭示了该类群复杂的倍性变化。研究以拉萨裂尻鱼为对象,完成了高质量的单倍型分型基因组组装,结果显示裂腹鱼的染色体数少于理论预期,主要由染色体融合而非丢失导致,且这些融合表现出不对称性。染色体融合推动了部分基因组区域向二倍体遗传模式转变,表明重新二倍化过程正在进行。研究还发现,基因丢失和表达分化现象与高原适应相关。本文揭示了裂腹鱼类群的共同自体多倍体化起源,并阐明了染色体融合和重新二倍化驱动基因组演化的机制。