

NP重磅 | 3个睡莲高质量T2T基因组
文章导读:
睡莲(Nymphaea)是最早分化的被子植物类群之一,保留了许多原始花器官特征。其花色与花香影响繁殖和生态适应,但相关起源和调控机制尚不清楚。作为早期分化类群,睡莲为研究被子植物起源及花植物早期进化提供了独特窗口。
海南大学研究团队于2026年5月6日在国际著名杂志Nature Plants上发表标题为“Water lily complete genomes illuminate the innovations of water lilies and early angiosperms”的研究性论文,构建了三种睡莲高质量 T2T 基因组并重建其系统发育,从基因组层面揭示了睡莲及被子植物早期演化的分子机制。

主要研究成果:
1.睡莲的优化系统发育研究
对223个睡莲样本进行系统发育分析,覆盖古热带、新热带、耐寒、澳洲和泛热带类型,并纳入若干外群物种。构建系统发育树结果显示,睡莲属主要分为两大分支:A组(日间开花)和B组(夜间开花),约在5900万年前分化。A组随后快速辐射,形成三个生态地理亚群:A1(热带睡莲)、A2(热带澳洲睡莲)和A3(耐寒睡莲),其分化时间与大陆漂移进程相对应。A1与A2进一步分化约发生在3700万年前。基于系统发育关系和基因组分析,筛选出蓝星睡莲N. colorata、侏儒睡莲Nymphaea thermarum和蓝睡莲Nymphaea caerulea作为高质量基因组测序候选物种,同时选择秘鲁睡莲Nymphaea sp. Peru Puerto Maldonado作为B组代表样本。
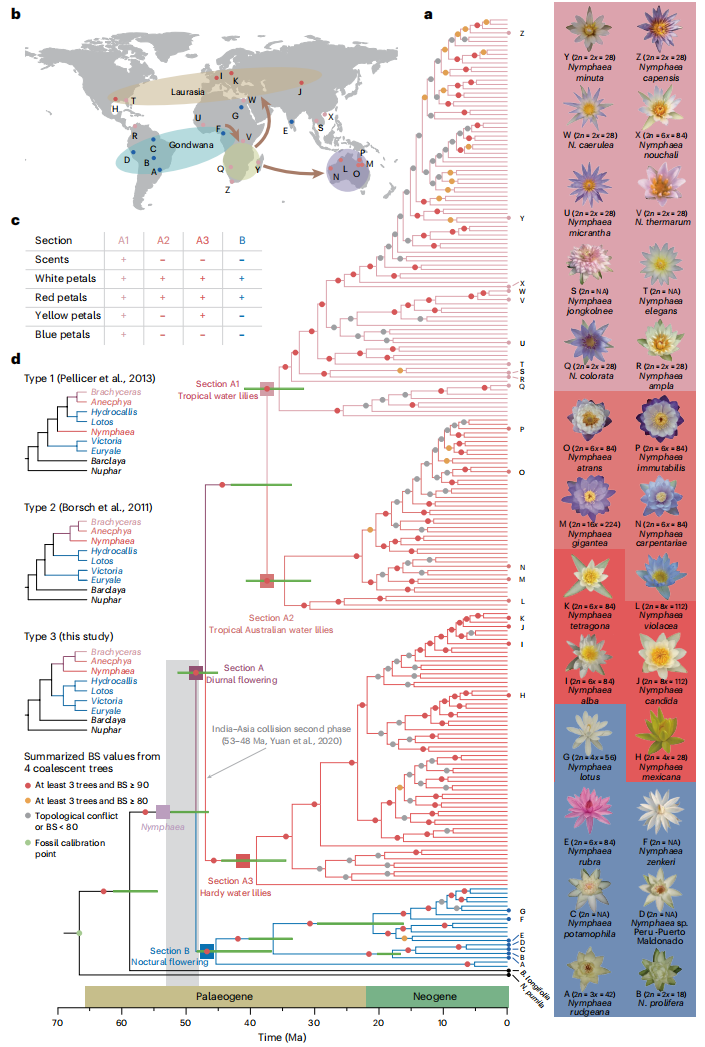
图1:睡莲属的系统发育与生物地理学分析
2.三种睡莲的T2T基因组组装
蓝星睡莲、侏儒睡莲和蓝睡莲三种睡莲基因组来自高度纯合自交系,采用PacBio HiFi、ONT超长读长及Hi-C数据组装,最终将三个基因组各自组装为14条拟染色体,实现无缺口组装。重复序列占比约35%~40%,主要为LTR反转录转座子。蛋白编码基因分别为21,239~28,911个,着丝粒和端粒几乎全部被鉴定,BUSCO完整性94.9%~97.1%。叶绿体基因组高度保守,长度约159.8 kb,基因组成一致,而线粒体基因组长度差异较大。此外,研究还完成了秘鲁睡莲的染色体级基因组,为比较分析提供参考基因组。
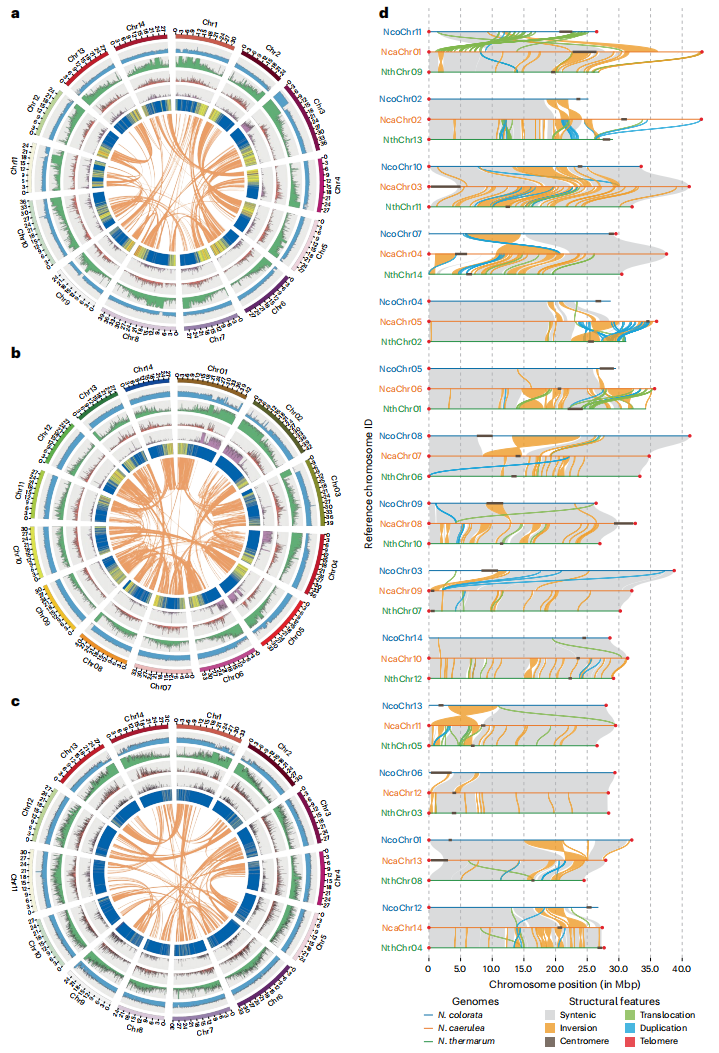
图2:基因组组装特征与共线性分析
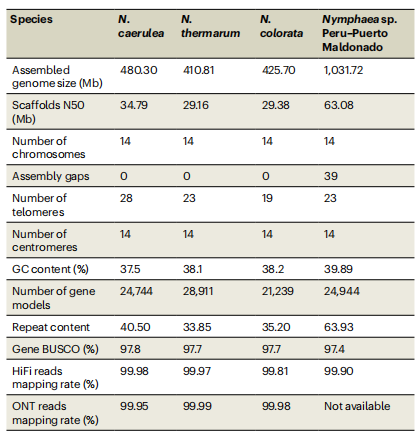
表1:睡莲基因组统计信息
3.睡莲目的染色体组进化
研究推断睡莲目祖先核型由13条祖先染色体组成,经历全基因组复制后形成26条祖先染色体,随后染色体裂变、丢失、端对端连接和嵌套融合等事件共同塑造了睡莲科(n = 14)和莼菜科(n = 15)的祖先核型。睡莲属祖先核型虽与睡莲科同为 n = 14,但仍发生6次裂变和6次融合。在属内,三种日间开花物种(A组)染色体结构较为稳定,而夜间开花物种(B组)相较祖先核型出现相互易位。相比之下,芡Euryale ferox染色体重排更为明显,与其谱系特异的全基因组三倍化历史相符。从睡莲目祖先到莼菜Brasenia schreberi的演化轨迹也表明,不同多倍化事件后染色体重排频率存在差异。
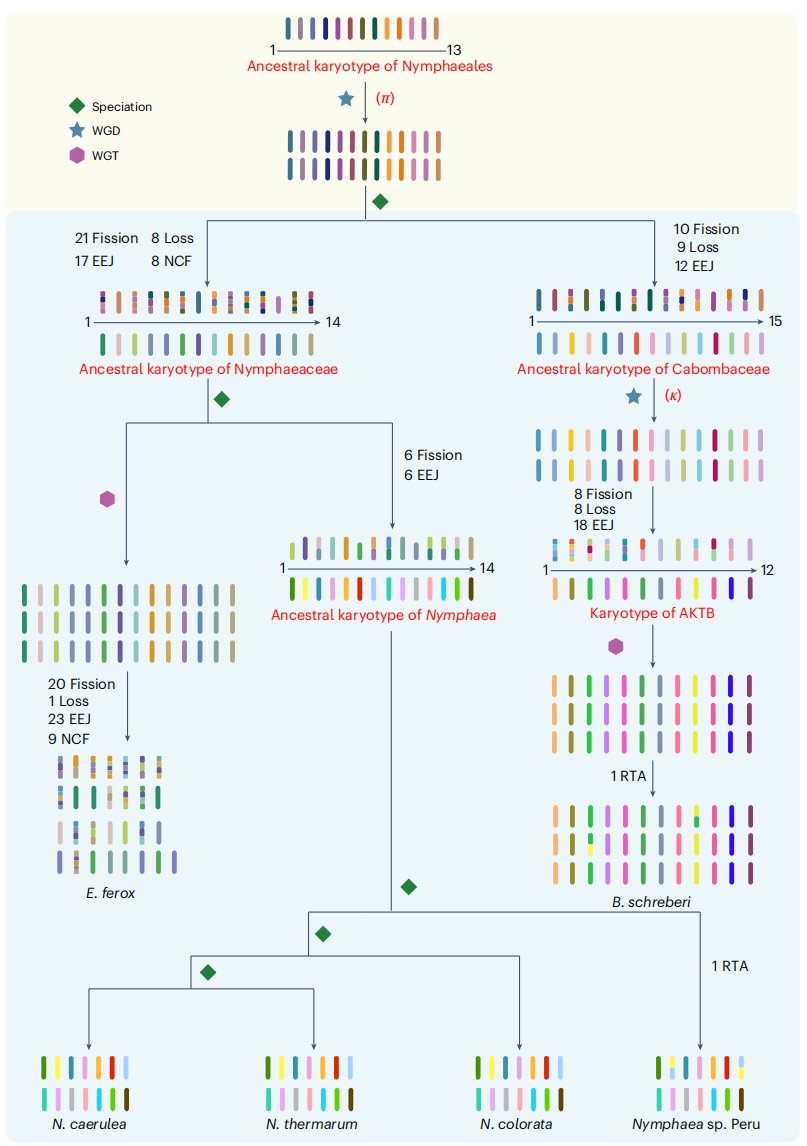
图3:睡莲目祖先核型重建与染色体演化
4.被子植物特异的直系群和基因家族
比较基因组分析显示,被子植物中有885个直系同源基因家族发生显著扩张,主要富集于DNA复制、基因表达调控、花粉壁组装等生物过程。进一步分析发现,被子植物特异基因家族还与木聚糖、果胶和木质素生物合成,以及低温胁迫响应等功能相关。结构域分析鉴定出213个被子植物特异蛋白结构域,其中含Pec_lyase_N与Pectate_lyase_4的双结构域果胶裂解酶在花粉中特异高表达,并具有保守α螺旋结构,其可能通过结构域叠加获得新功能。功能上,该类果胶裂解酶可能通过定向降解花柱组织中富含果胶的中胶层,促进花粉管穿透并到达胚珠,同时维持周围细胞结构完整。
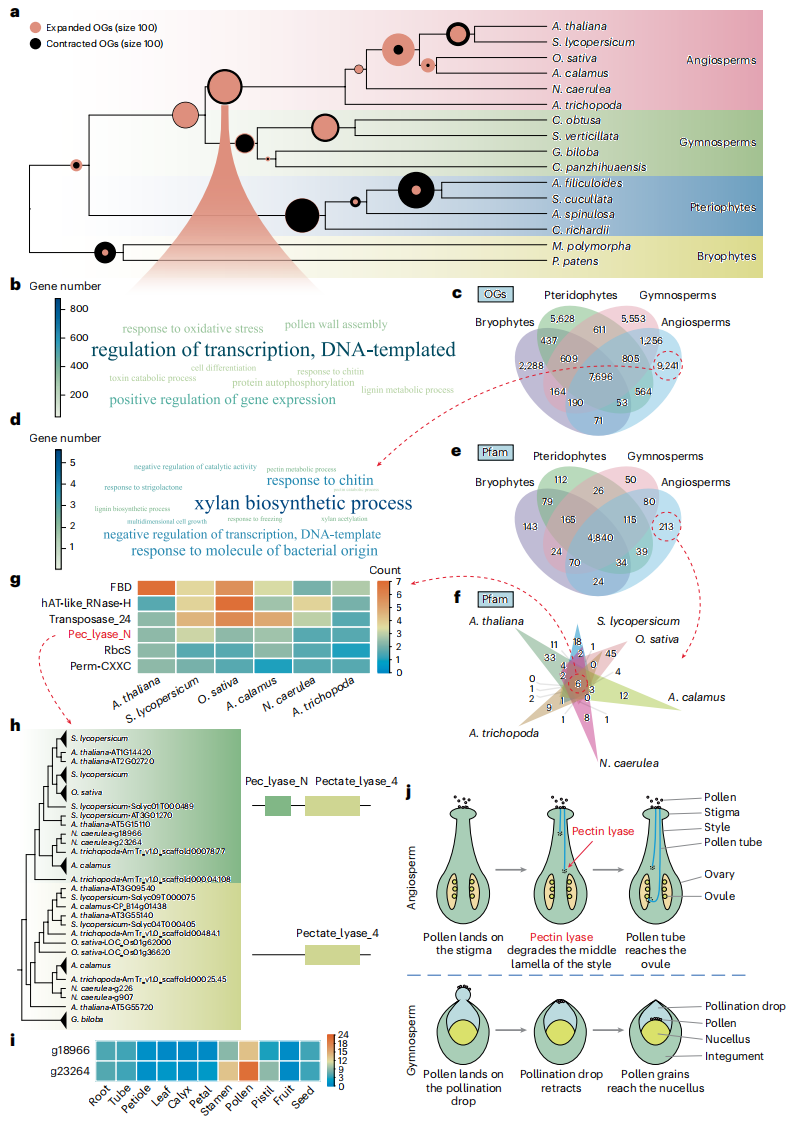
图4:被子植物特异基因获得揭示促进授粉的果胶裂解酶
5.睡莲在适应水生环境过程中基因的丢失
睡莲目植物作为全水生植物,其进化适应主要体现在多个基因家族的丢失:水的浮力减少了对机械支撑的需求,导致许多与细胞壁发育相关的基因丢失;水生环境中陆生致病微生物无法生存,引起相关抗性基因丢失;此外,陆生植物中用于防御捕食者的毒素代谢途径在水生环境中变得冗余,导致相关代谢基因减少。这些基因丢失反映了睡莲在水生环境中形态与生理特征的特化进化,为理解水生植物的适应机制提供了基因组学依据。
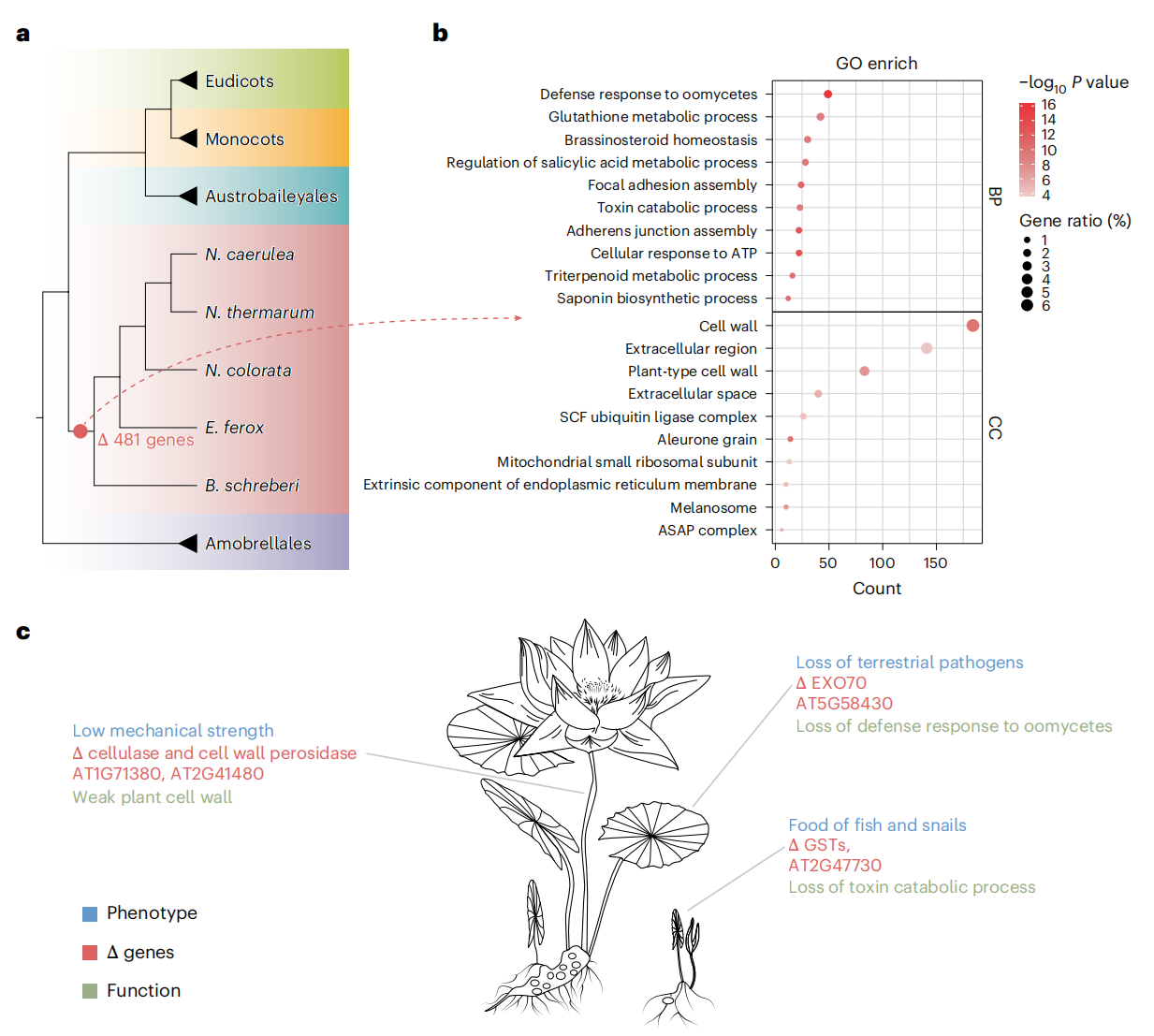
图5:睡莲中的基因丢失揭示其水生生活适应相关的候选基因
6.花色素调控及功能验证
通过代谢组学发现,蓝星睡莲不同花色品种中主要积累的花青素为矢车菊素-3-O-半乳糖苷,含量随花色差异显著。转录组加权共表达网络分析显示,MYB 转录因子主导与花青素生物合成相关的核心调控模块。功能验证表明,蓝星睡莲的NcolMYB75-like在转基因拟南芥中过表达可显著促进花青素积累,并上调结构基因表达。进一步实验显示,NcolMYB75-like能结合并激活NcolF3H-like启动子,从而调控花青素合成。
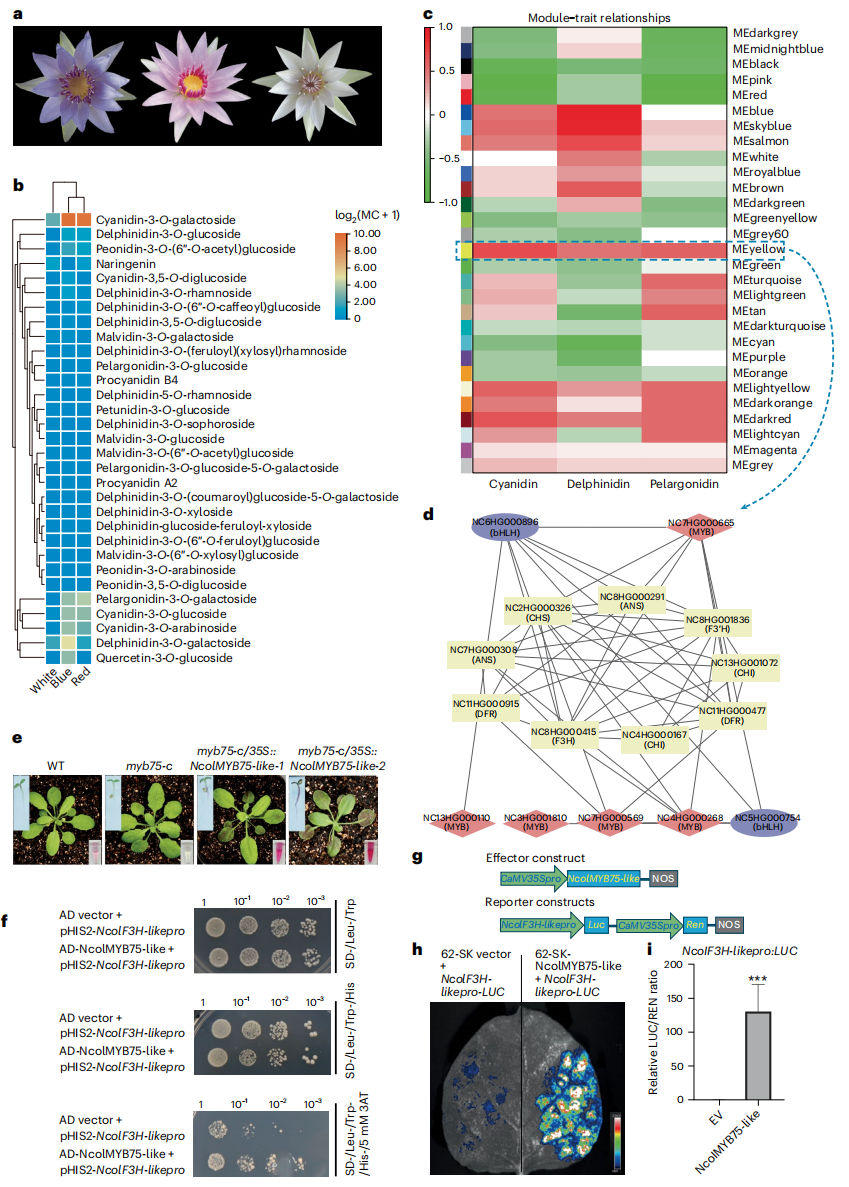
图6:不同花色蓝星睡莲品种的代谢谱与遗传解析
7.睡莲花香组成与OMT酶功能研究
研究显示,睡莲花香在物种间存在显著差异:蓝星睡莲以p-茴香醛和p-茴香醇为主,蓝睡莲含八种甲基苯酚,而侏儒睡莲不含甲基苯酚,仅释放苯丙素衍生物。甲基苯酚在组织来源和释放时间上呈物种特异性,反映了不同的传粉者吸引策略。基因组分析表明,睡莲科OMT家族相较基干被子植物显著扩张,并形成谱系特异分支。功能验证显示,NcOMT1与NcaOMT1/2/3 在各自物种中具有特异甲基转移活性,将前体转化为物种特异花香成分。
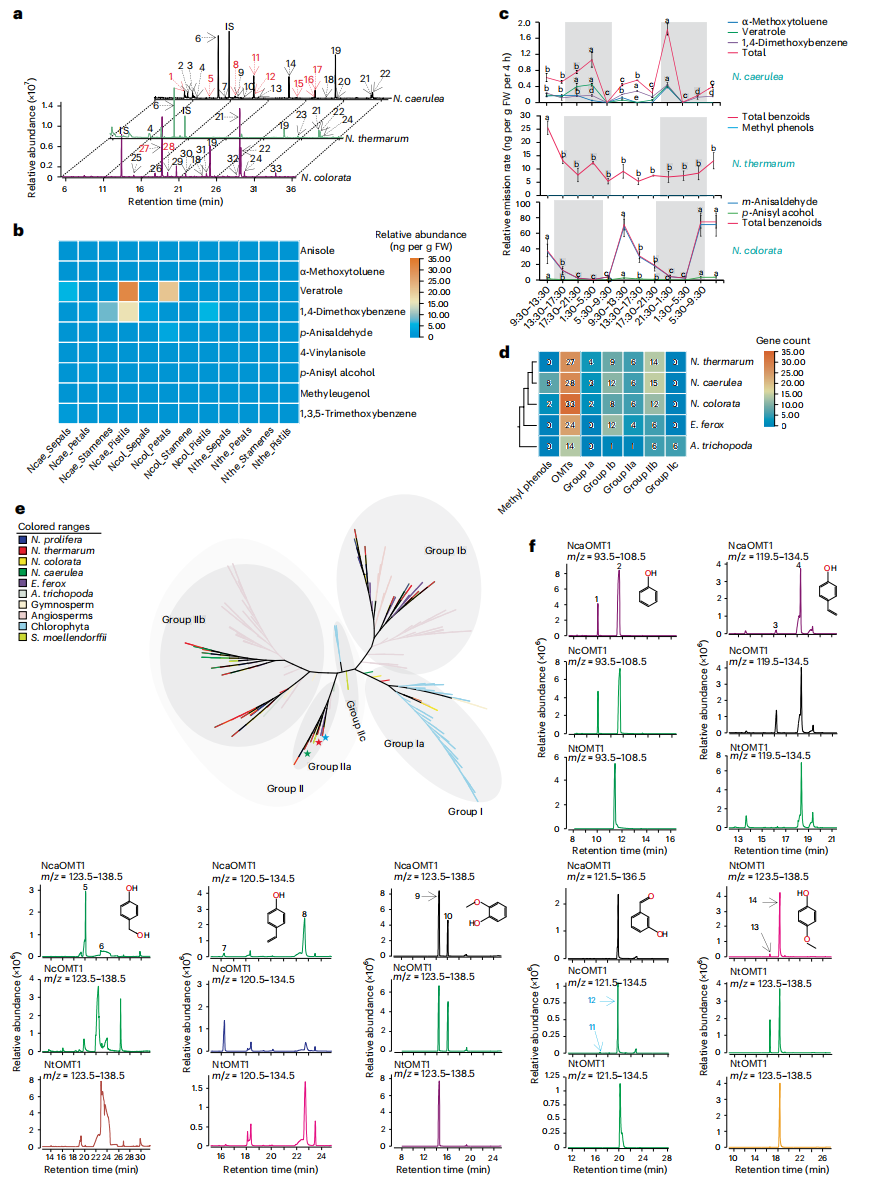
图7:睡莲属物种的花香化合物及 OMT 系统发育树
文章小结
本研究构建了三种睡莲(N. colorata、N. caerulea、N. thermarum)的高质量T2T基因组,并结合广泛取样重建了睡莲属系统发育,明确其主要分支及演化历史。通过比较基因组学和多组学分析,研究从授粉机制、花色形成、花香挥发物合成和水生适应等方面,揭示了睡莲及基干被子植物早期演化创新的分子基础。本研究结果不仅加深了对睡莲属基因组演化和生态适应的理解,也为花卉性状改良、种质资源利用和水生植物保护提供了重要基因组资源。