

Cell重磅 | 基因组揭示深海生物超强耐饿机制
深海环境长期处于高压、低温和食物匮乏状态,生物面临严峻的能量获取挑战。尽管如此,深海巨型动物普遍表现出显著的巨型化特征,并具备极强的耐饥能力。它们如何在有限能量供给下维持生存和生长,一直是深海生物适应性进化研究的重要问题。
中国科学院海洋研究所等单位于2026年6月5日联合在Cell发表标题为“Deep-sea megafauna co-opts microbial energy metabolism genes to withstand ultra-long starvation”的研究论文,揭示了深海巨型等足类动物应对长期食物匮乏的新机制。研究发现,其祖先通过水平基因转移获得了来源于微生物的能量代谢基因,并在后续进化过程中通过表观遗传调控实现功能优化,从而形成低代谢、高耐饥的生存策略,破解了巨大体型与极端能源匮乏之间的生存矛盾。

主要研究成果
1.深海超巨型等足类动物的能量悖论
作者比较了潮间带等足类 Cirolana isopus、道氏深水虱Bathynomus doederleini 和詹姆斯深水虱Bathynomus jamesi 的形态与生理特征,发现其呈现显著的“越深越大”规律,即随着栖息深度增加,体型和胃容量同步增大。为探究这一适应现象背后的遗传基础,作者进一步完成了道氏深水虱的染色体水平基因组组装(~ 4.35 Gb),并与此前发表的詹姆斯深水虱基因组进行比较分析。结果表明,深海物种具有更强的食物储存与营养利用能力、更高的能量储备以及更低的基础代谢水平;同时,其消化吸收相关基因家族显著扩张,而能量产生相关的氧化磷酸化核心基因家族发生收缩。
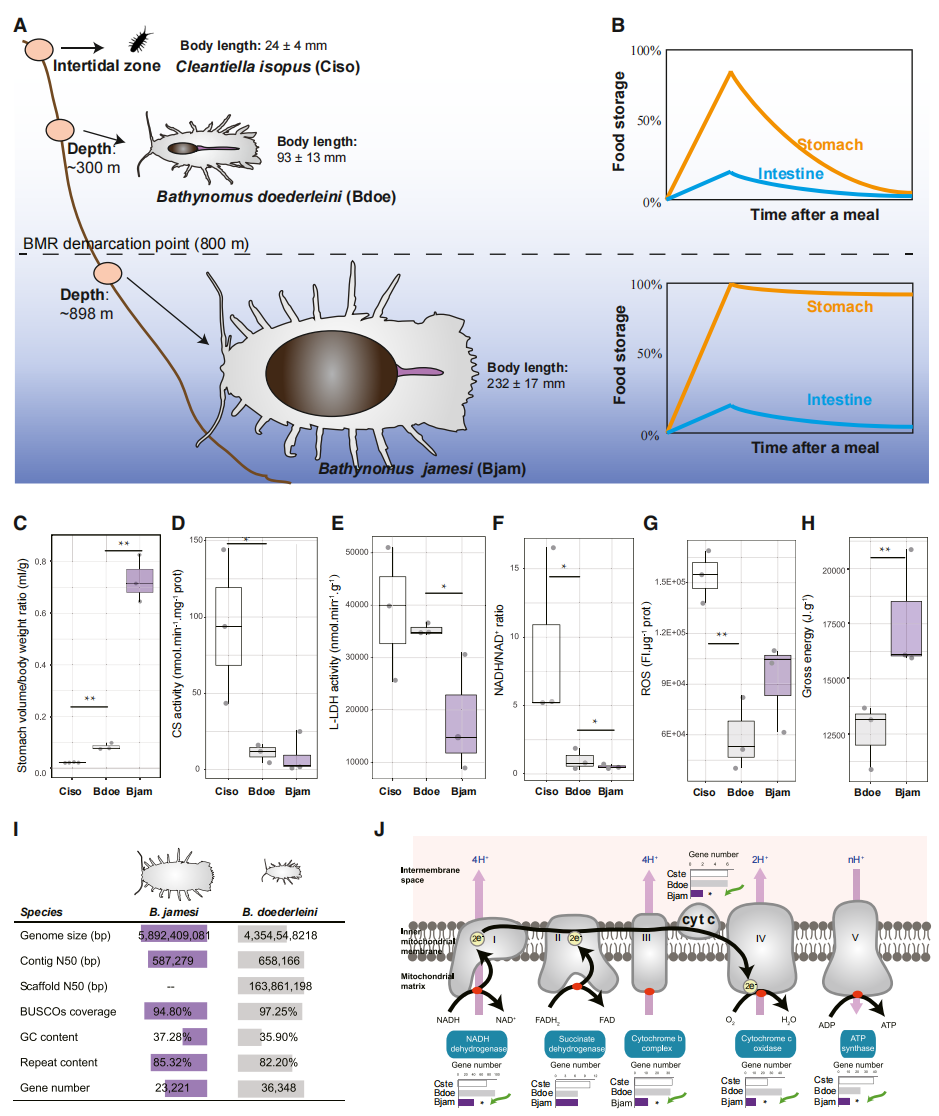
图1:两种深海等足类动物在胃部储食能力和基础代谢率上的差异
2.深海胃微生物群适应性演化
为探究微生物群在深海等足类耐饥适应中的作用,作者对詹姆斯深水虱和道氏深水虱的胃肠微生物组进行了宏基因组分析。结果发现,尽管两者肠道微生物组成相似,但胃部微生物群存在显著差异。较浅海种道氏深水虱富集能够促进食物消化和能量利用的厚壁菌门和柔膜菌门微生物,而深海超巨型种詹姆斯深水虱则富集具有特殊代谢能力的衣原体类群。进一步分析表明,詹姆斯深水虱胃部微生物组缺乏复杂碳水化合物降解相关酶系,整体消化和能量利用能力较弱,却富集与转录调控和组蛋白乙酰化相关基因,这说明深海超巨型等足类形成了与长期储能和低代谢状态相适应的特异性胃部微生物群落。
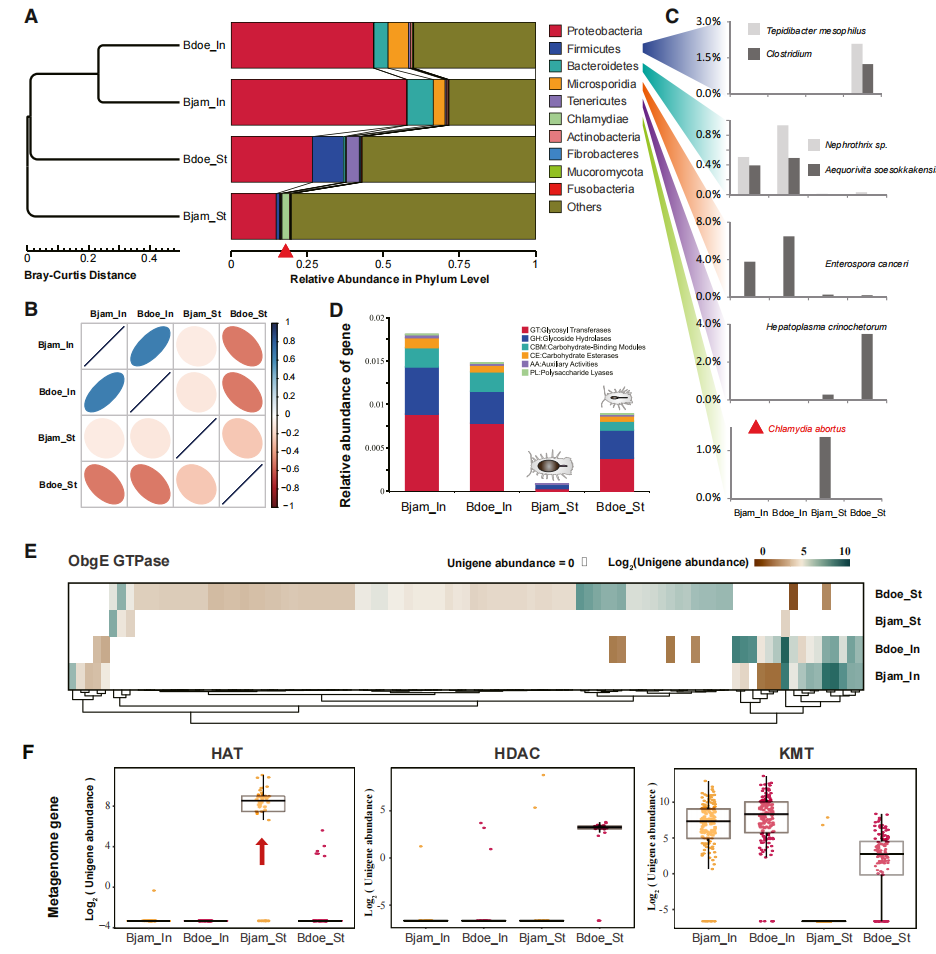
图2:宿主与内共生微生物的能量代谢协同进化
3.深海等足类的水平转移基因
为探究深海等足类极端耐饥能力的遗传基础,作者对两种深海大王具足虫基因组进行了系统筛查,共鉴定出ND1、NS1和RdRP等3类来源于细菌或病毒的水平转移基因。其中,ND1与细菌来源的NADH脱氢酶具有显著同源性。结合序列特征、基因结构、共线性分析及PCR验证,研究证实这些外源基因已稳定整合至宿主染色体。系统发育分析进一步表明,ND1的水平转移事件发生于两种大王具足虫分化之前,距今约1600万年。且这些水平转移基因在长期进化过程中逐渐获得与宿主相似的密码子使用偏好,并部分演化出真核生物特有的多外显子结构,表明其已完成深度基因组驯化并融入宿主遗传体系。
4.ND1基因的适应性扩增与驯化
在鉴定出来源于微生物的水平转移基因后,作者进一步分析了其在宿主基因组中的演化命运。研究发现,ND1在进入深海等足类基因组后经历了显著的拷贝数扩增,并在两种大王具足虫分化后分别发生独立复制,表现出平行演化特征。与此同时,ND1还突破了水平转移基因通常低表达的规律,在两种深海物种中均呈现异常高的转录水平,其中多个拷贝位列全基因组表达量最高的1%。进一步分析表明,ND1形成了区别于宿主旁系同源基因的密码子使用偏好性,其可能经历了适应性选择,从而促进了基因的超高表达。
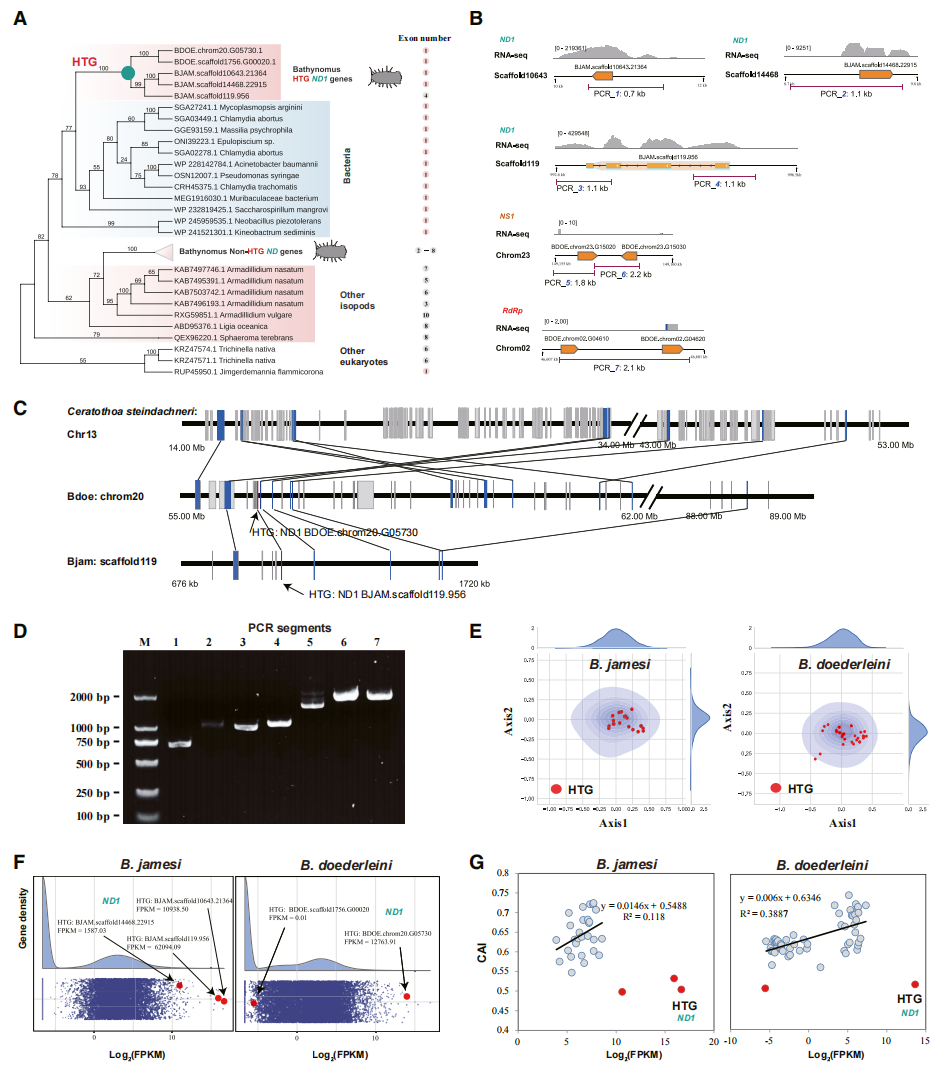
图3:深海等足类基因组中的古老水平转移基因
5.组蛋白乙酰化增强
进一步分析两种深海大王具足虫的组蛋白及其修饰酶基因家族,结果发现,深海等足类的组蛋白核心基因(H1、H2A、H2B和H4)发生显著收缩,而组蛋白乙酰转移酶(HAT)和赖氨酸甲基转移酶(KMT)等修饰酶基因则明显扩张,呈现出组蛋白结构蛋白与调控酶“此消彼长”的进化模式。其中,HAT在詹姆斯深水虱胃组织中受到强烈正向选择,不仅发生串联扩增,还表现出胃组织特异性高表达。与此同时,其胃部共生微生物组中也富集了HAT相关基因,表明宿主与微生物协同强化了组蛋白乙酰化调控体系。
6.乙酰化驱动的ND1高表达
为揭示ND1超高表达的调控机制,作者进一步分析了深海大王具足虫胃组织中的组蛋白乙酰化修饰。结果发现,深海巨型种詹姆斯深水虱具有显著增强的组蛋白乙酰化水平,在转录起始位点附近形成更强的H3K9ac富集信号。进一步分析表明,多个高表达ND1拷贝的启动子区域均存在明显的H3K9ac峰,而低表达拷贝则缺乏相应修饰,呈现出乙酰化水平与表达强度高度一致的对应关系。结合前文发现的HAT基因扩张及高表达现象,作者提出深海等足类演化出强化的组蛋白乙酰化调控体系,通过特异性激活外源基因ND1,实现其超高表达,并最终促进低代谢和长期耐饥表型的形成。
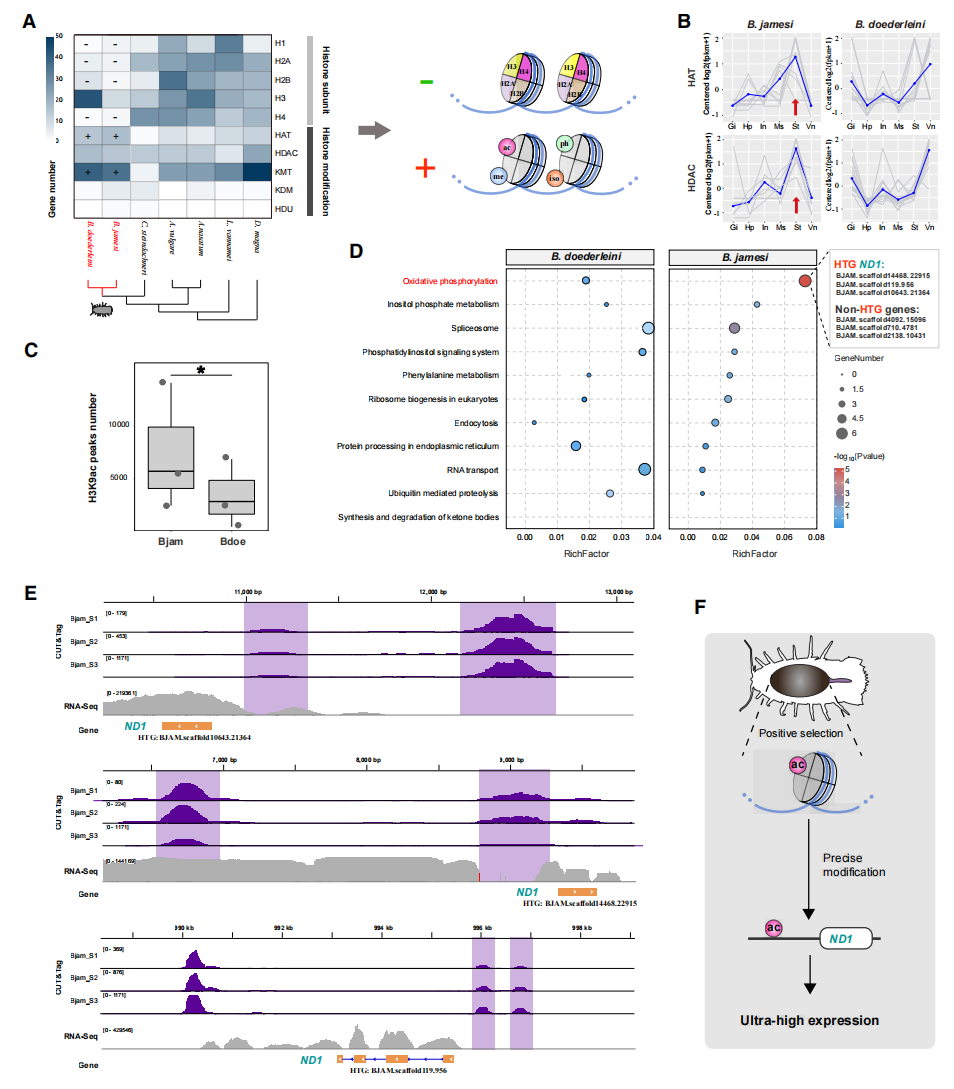
图4:组蛋白乙酰化特异性驱动ND1超高表达
7.ND1驱动低代谢耐饥
为验证ND1是否是深海等足类超强耐饥能力的关键因子,作者在斑马鱼、293T细胞和秀丽隐杆线虫中开展了跨物种功能验证。结果发现,ND1能够显著重塑宿主能量代谢状态,但其作用具有明显的环境依赖性:在常温条件下,ND1促进能量产生相关基因表达并增强代谢活动;而在低温诱导的低基础代谢状态下,则进一步降低氧耗率、活性氧水平及NADH/NAD⁺比值,抑制线粒体能量代谢,从而延缓能量储备消耗并显著提高耐饥能力。结合不同模式生物中一致的表型结果,作者提出,ND1并非简单的代谢促进因子或抑制因子,而是一个能够根据环境状态动态调节能量分配的“能量权衡器”。在低代谢环境下,ND1通过进一步压低能量消耗,实现生长与生存之间的能量再分配,从而帮助宿主以更低成本维持长期存活。
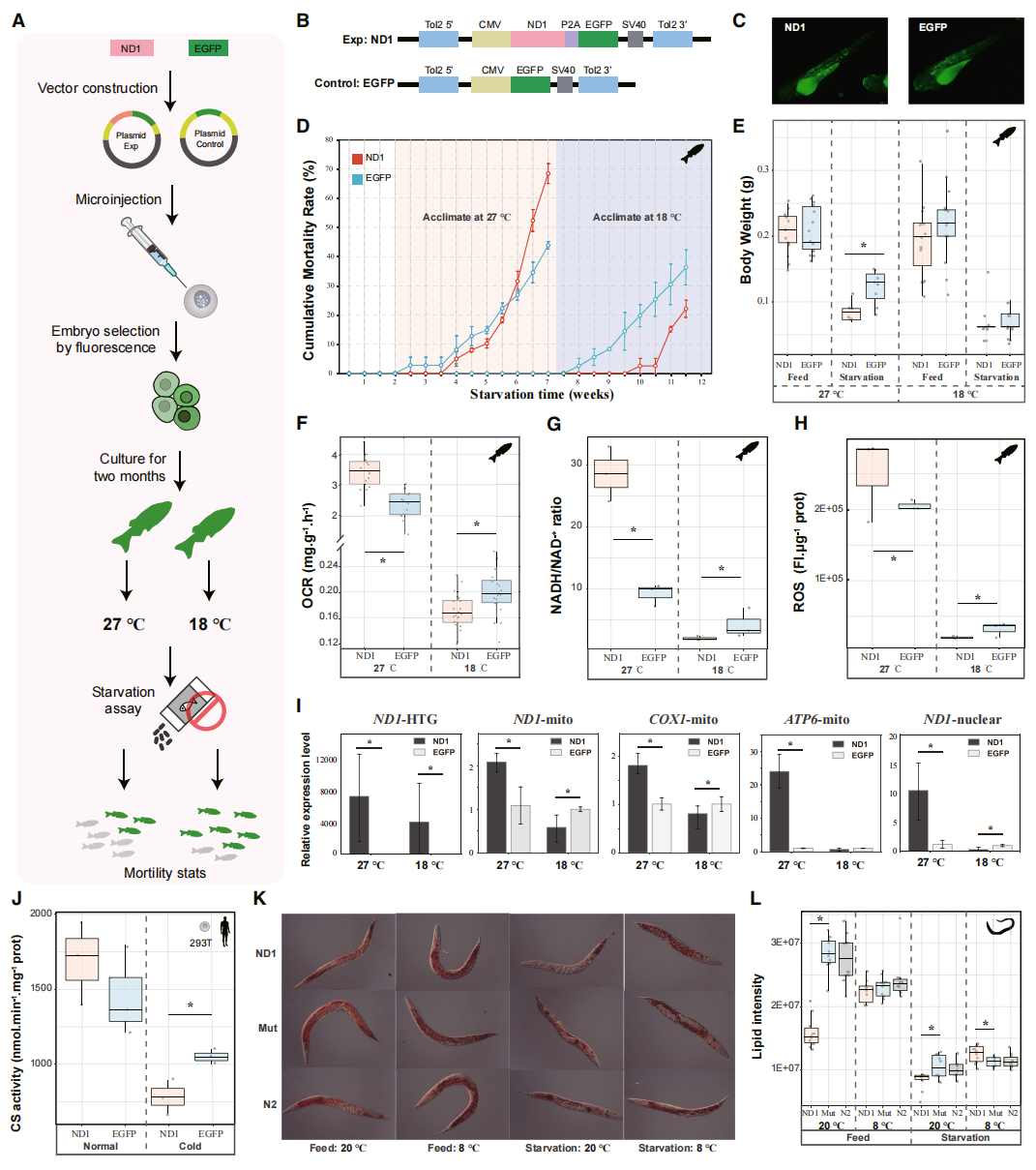
图5:ND1水平转移基因在低代谢状态下延长耐饥时间
文章小结
本研究中,作者通过比较不同深度分布的大王具足虫,结合基因组学、宏基因组学、表观遗传学和跨物种功能实验发现,深海超巨型等足类能够通过扩大胃容量、降低基础代谢率和优化能量利用来适应长期食物匮乏环境。进一步研究表明,其祖先通过水平基因转移获得了来源于微生物的能量代谢相关基因ND1,并在后续进化过程中经历基因扩增和组蛋白乙酰化驱动的超高表达。功能实验显示,ND1能够根据环境状态动态调控能量代谢,在低代谢条件下抑制能量消耗、提高耐饥能力,从而帮助深海巨型动物实现长期生存。